Cached at:
06/08/26, 03:34 AM
# Transgenic hookworm secretes anti-tetrodotoxin human single chain antibody
Source: [https://www.nature.com/articles/s41467-026-73447-9?error=cookies_not_supported&code=1270f4db-8d1f-4c44-88bf-78d0133011e7](https://www.nature.com/articles/s41467-026-73447-9?error=cookies_not_supported&code=1270f4db-8d1f-4c44-88bf-78d0133011e7)
## Introduction
Parenteral administration delivers medications and biologics directly into the body, bypassing the gastrointestinal \(GI\) tract to ensure rapid and effective absorption\. Conventional methods \(such as intravenous, subcutaneous, intramuscular, and intradermal injections\) are used for therapies requiring precise dosing or fast onset\. However, novel delivery systems are being developed to improve the efficiency, stability, and targeting of biologics, with growing interest in biological organisms, e\.g\., parasite\-based delivery of genes and proteins to neurons[1](https://www.nature.com/articles/s41467-026-73447-9#ref-CR1)\. Similarly, hookworms can be explored as an innovative pharmaceutical biofactory platform for both drug production and direct delivery to the human gut[2](https://www.nature.com/articles/s41467-026-73447-9#ref-CR2)\. Hookworms have evolved to survive for years within the human host while minimally disrupting host homeostasis[3](https://www.nature.com/articles/s41467-026-73447-9#ref-CR3), and controlled human infections with hookworms are safe and well\-tolerated in clinical settings[4](https://www.nature.com/articles/s41467-026-73447-9?error=cookies_not_supported&code=1270f4db-8d1f-4c44-88bf-78d0133011e7#ref-CR4),[5](https://www.nature.com/articles/s41467-026-73447-9?error=cookies_not_supported&code=1270f4db-8d1f-4c44-88bf-78d0133011e7#ref-CR5),[6](https://www.nature.com/articles/s41467-026-73447-9#ref-CR6), bolstering their potential for utility as pharmaceutical biofactories[2](https://www.nature.com/articles/s41467-026-73447-9#ref-CR2)\. This opens an alternative approach for in vivo constitutive hookworm\-based therapeutic delivery, and given that different helminths occupy diverse tissue niches, defined helminth species can deliver transgene\-derived products to specific organs and tissues\.
The development of tools for functional genomics in parasitic nematodes has faced significant challenges\. RNA interference \(RNAi\), one of the primary tools for gene knockdown, has shown variable efficacy and is limited to only a few parasitic nematode species[7](https://www.nature.com/articles/s41467-026-73447-9#ref-CR7),[8](https://www.nature.com/articles/s41467-026-73447-9#ref-CR8)\. Clustered Regularly Interspaced Short Palindromic Repeats/ CRISPR\-associated proteins \(CRISPR/Cas9\), a revolutionary technology for genome editing, has been adapted for use in a small number of parasitic nematodes, prominently*Strongyloides stercoralis*and*S\. ratti*\(e\.g\., ref\.[9](https://www.nature.com/articles/s41467-026-73447-9#ref-CR9)\)\. In such cases, microinjection protocols developed for the free\-living*Caenorhabditis elegans*were used due to the ability of these parasitic nematode species to complete the whole life cycle outside of the mammalian host[9](https://www.nature.com/articles/s41467-026-73447-9#ref-CR9)\. A single report used a*piggyBac*\-mediated transfection system for CRISPR\-based insertion and resulted in a ~3% transfection rate for the microfilariae stage of*Brugia malayi*, making screening of transgenic adults difficult after completing the life cycle[10](https://www.nature.com/articles/s41467-026-73447-9#ref-CR10)\. This approach may also lead to multi\-copy insertions\. Because of the complexities of the life cycles of parasitic nematodes, coupled with their large, complex, and poorly annotated and characterized genomes, CRISPR/Cas\-based programmed knock\-out \(KO\) and knock\-in \(KI\) transgenesis has not been achieved in many species\. Significant hurdles include a lack of identified genomic safe harbors \(GSHs, specific loci in the genome where introduced DNA can be inserted without disrupting vital genes or regulatory elements[11](https://www.nature.com/articles/s41467-026-73447-9#ref-CR11)\) that can facilitate targeted transgene insertion, which is challenging given the lack of data on the availability and accessibility of chromatin in these organisms\. Moreover, due to the thick cuticle of parasitic nematodes \(compared to*C\. elegans*\), which protects the worm from the host’s immune system and environmental stress, more effective delivery methods to facilitate transfection need to be identified to evaluate CRISPR/Cas9 systems that can function reliably in a variety of nematode species[12](https://www.nature.com/articles/s41467-026-73447-9#ref-CR12)\. Additional obstacles for pharmaceutical biofactory development include limited germ\-line access for heritable transgenesis and the substantial knowledge gap around parasitic nematode secretion systems\. As a first step towards developing hookworms as living pharmaceutical factories capable of producing and delivering therapeutic proteins directly inside the host, a gain\-of\-function transgenesis protocol must be established to enable stable expression of therapeutic transgenes within the hookworm’s genome\.
Here, we report on methodological, technical, and conceptual advances, demonstrating successful bioengineering of a human hookworm,*Ancylostoma ceylanicum*, to produce and secrete a human single\-chain antibody, s16\-HuScFv, that neutralizes tetrodotoxin \(TTX\)[13](https://www.nature.com/articles/s41467-026-73447-9#ref-CR13)\. We first identify and prioritize two GSHs and confirm successful chromosomal cleavage at CRISPR\-targeted sites in the hookworm genome using next\-generation sequencing\. Next, we perform CRISPR\-programmed knock\-in of the*s16\-HuScFv*transgene into one of the GSHs in the genome of the egg stage of*A\. ceylanicum*\. To confirm successful transgenesis, a lack of disruption in surrounding gene expression, and transmission through the life cycle, we present results from genomic sequencing, qRT\-PCR, and RNA\-seq analysis\. Finally, we demonstrate TTX neutralization mediated by circulating s16\-HuScFv in the blood of hamsters infected with the transgenic hookworms\. This advancement represents a critical step towards the development of a transgenic human hookworm pharmaceutical biofactory platform with the potential to continuously, safely, and effectively deliver biologics in situ within patients[2](https://www.nature.com/articles/s41467-026-73447-9#ref-CR2)\.
## Results
### Identification, prioritization, and experimental validation of Genome Safe Harbor \(GSH\) regions
At the outset, we identified GSH regions and prioritized two \(GSH1 and GSH2\) for insertion of the s16\-HuScFv transgene cassette and other corresponding construct elements \(Figs\.[1](https://www.nature.com/articles/s41467-026-73447-9#Fig1)a,[b](https://www.nature.com/articles/s41467-026-73447-9#Fig1)\) in the hookworm genome, based on results from a multi\-omics driven approach utilizing available*A\. ceylanicum*data \(see Methods\)\. Putative 5’ upstream promoter sequences associated with genes highly expressed across the*A\. ceylanicum*life cycle \(19 RNA\-seq datasets[14](https://www.nature.com/articles/s41467-026-73447-9#ref-CR14),[15](https://www.nature.com/articles/s41467-026-73447-9#ref-CR15)\) were identified using STREME[16](https://www.nature.com/articles/s41467-026-73447-9#ref-CR16)\. This analysis utilized a database of functional annotations, RNAseq based gene expression[14](https://www.nature.com/articles/s41467-026-73447-9#ref-CR14),[15](https://www.nature.com/articles/s41467-026-73447-9#ref-CR15)and mass spectrometry\-derived excretory/secretory protein \(ESP\) proteomics data[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)for all 18,776 genes encoded in the*A\. ceylanicum*genome \(Supplementary Data[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM3); Supplementary Fig\.[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. Two genes were identified with high expression, evidence of protein secretion, and putative promoter sequences associated with the top 1% \(GSH1\) or the top 2% \(GSH2\) of expressed genes, as detailed below\. This approach was utilized to identify genes that are consistently accessible across the life cycle, associated with upstream promoter sequences resulting in high gene expression and putatively secreted from secretory tissues contributing to ESPs\.
**Fig\. 1: Graphical representation of the double stranded donor construct and the genome safe harbor \(GSH2\) region\.**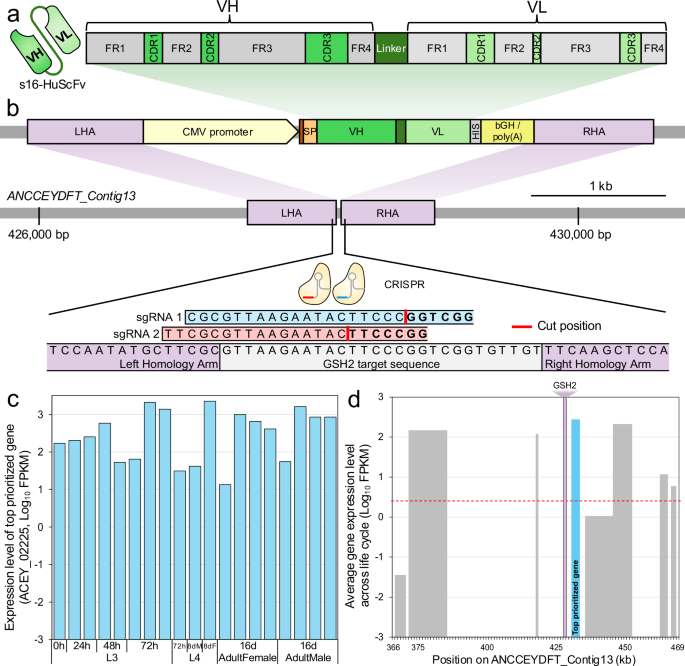The alternative text for this image may have been generated using AI\.
[Full size image](https://www.nature.com/articles/s41467-026-73447-9/figures/1)
GSH2 was identified upstream of the prioritized target gene*maker\-ANCCEYDFT\_Contig13\-pred\_gff\_snap\-gene\-4\.9*/*ACEY\_002225*\.**a**Structural domains of the s16\-HuScFv protein, as defined in Chulanetra et al, 2012[13](https://www.nature.com/articles/s41467-026-73447-9#ref-CR13)\. Individual immunoglobulin frameworks \(FRs\) and complementarity\-determining regions \(CDRs\) are indicated for the Variable Heavy \(VH\) and Variable Light \(VL\) components of s16\-HuScFv, bound by a linker sequence\.**b**The genetic components of the double\-stranded donor construct and its insertion into the*A\. ceylanicum*genome at the GSH region of the “*ANCCEYDFT\_contig13*” contig\. The construct was engineered by cloning the human single\-chain antibody fragment \(s16\-HuScFv\) into pCDNA3\.1\(\+\) backbone \(at NotI restriction site\), with a CMV promoter, a polyhistidine \(HIS\) tag, and bovine growth hormone polyadenylation termination sequence \(bGH/poly\(A\)\)\. The donor construct was flanked at its termini with homology arms of ~600 bp each \(HA\)\. The positions of the s16\-HuScFv Variable Heavy \(VH\) and Variable Light \(VL\) chain regions and the linker sequence are shown in green\. The left homology arm \(LHA\) is situated at the position of two overlapping single guide RNAs \(sgRNAs\), sgRNA 1 \(ranked 5th by CHOPCHOP\) and sgRNA 2 \(ranked 13th\)\. The terminal residues of sgRNA sequences are the CGG protospacer adjacent motif\.**c**The average gene expression level of*ACEY\_002225*\(log\-scale FPKM\) across the*A\. ceylanicum*life cycle, including larval \(L3, L4\) and adult stages\. The dashed red line indicates the average expression level of all genes across all stages\.**d**The gene expression levels of genes across the GSH region on the*ANCCEYDFT\_Contig13*contig of the genome assembly\. X\-axis positions indicate gene coordinates, and Y\-axis values represent gene expression levels\. The prioritized target gene*ACEY\_002225*is indicated in blue; the dashed red line indicates the average expression level of all genes across all stages\. The purple shaded area running vertically up the plot indicates the target “GSH2” region\. There is 12,966 bp of sequence preceding the target gene on the genome assembly\. Source data are provided as a Source Data file\.
For the first putative GSH, we identified an enriched sequence motif among the 5’ upstream regions of the top 1% of all expressed*A\. ceylanicum*genes across the life cycle \(CACTCGTAA;*P*= 0\.0016; STREME[16](https://www.nature.com/articles/s41467-026-73447-9#ref-CR16)binomial distribution test; Supplementary Data[2](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\), annotated[18](https://www.nature.com/articles/s41467-026-73447-9#ref-CR18)as a*ceh\-22*binding motif \(*P*= 0\.0026\)\. Among genes with this motif, we prioritized*ACEY\_000382*, a protein disulfide\-isomerase that is more highly expressed than 99\.83% of all*A\. ceylanicum*genes, contains a signal peptide for secretion[19](https://www.nature.com/articles/s41467-026-73447-9#ref-CR19), is orthologous to a highly abundant ESP in*A\. caninum*and*Necator americanus*hookworms[20](https://www.nature.com/articles/s41467-026-73447-9#ref-CR20),[21](https://www.nature.com/articles/s41467-026-73447-9#ref-CR21), and was detected in the ESPs of male \(25 peptides\) and female \(21 peptides\) adult*A\. ceylanicum*[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\. A protein disulfide\-isomerase was also among the most abundant proteins in the secretomes of*Schistosoma mansoni*[22](https://www.nature.com/articles/s41467-026-73447-9#ref-CR22)and*S\. japonicum*[23](https://www.nature.com/articles/s41467-026-73447-9#ref-CR23), and in the root knot nematode*Meloidogyne gramini*cola[24](https://www.nature.com/articles/s41467-026-73447-9#ref-CR24), one is secreted from the esophageal gland, where it likely interacts with host extracellular matrix components[22](https://www.nature.com/articles/s41467-026-73447-9#ref-CR22)\.*ACEY\_000382*is flanked by several other highly expressed genes across the life cycle, with the running average of expression level for five consecutive genes being the second highest in the genome \(Supplementary Fig\.[2a](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. The first prioritized genomic sequence for gRNA design was the intergenic region more than 1,346 bp upstream of*ACEY\_000382*, to avoid potentially interrupting any putative promoter regions\. We termed this region “GSH1”\.
The same approach was used for the second GSH using the top 2% of all expressed genes, identifying a 62 bp palindromic motif sequence associated with the top\-enriched[16](https://www.nature.com/articles/s41467-026-73447-9#ref-CR16)fkh\-2 motif[18](https://www.nature.com/articles/s41467-026-73447-9#ref-CR18),[25](https://www.nature.com/articles/s41467-026-73447-9#ref-CR25)\(GGTAAACGTGTTTACGCCCGTGTTAACGATGGGTAAATTTGTGTGGTGTAAACACGT\-TTACC; Supplementary Fig\.[1a](https://www.nature.com/articles/s41467-026-73447-9#MOESM1); See methods\)\. Among the genes with this upstream motif, we prioritized*ACEY\_002225*\(Immunogenic protein 3\) which is expressed more highly than 97\.8% of all*A\. ceylanicum*genes \(Fig\.[1c](https://www.nature.com/articles/s41467-026-73447-9#Fig1)\), contains a signal peptide for secretion, is an ortholog of an ESP in*A\. caninum*, and was detected in the ESPs of male \(6 peptides\) and female \(8 peptides\) adult*A\. ceylanicum*[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\(Supplementary Fig\.[1b](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\.*ACEY\_002225*, on contig13, is surrounded by several other highly expressed genes \(with the highest running average on the contig; Supplementary Fig\.[2b](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. The second prioritized genomic sequence used to design gRNA sequences was the intergenic region more than 1591 bp upstream of*ACEY\_002225*\(Fig\.[1d](https://www.nature.com/articles/s41467-026-73447-9#Fig1); Supplementary Fig\.[1c](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. We avoided the first 1591 bp, aiming not to interrupt any of the putative enriched promoter sequences \(provided in Supplementary Data[3](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\)\. We termed this region “GSH2”\.
Both GSH regions, surrounded by highly expressed genes over the life cycle, offer advantages for targeted gene insertion\. These regions exhibit high transcriptional activity, which reduces the risk of transgene silencing, ensuring that transgenes remain active and express their cargo proteins consistently over time and that surrounding chromatin is open and accessible for efficient transgene integration\. For each set of five genes per GSH in*A\. ceylanicum*\(two upstream, the GSH target gene, two downstream\), the human hookworm*N\. americanus*orthologs were all reciprocal BLAST hits \(i\.e\., the same protein pair was identified when searching from*A\. ceylanicum*to*N\. americanus*, and from*N\. americanus*to*A\. ceylanicum*\), and they were also in sequential order on the*N\. americanus*genome assembly, with the same strand orientations relative to the GSH target gene ortholog\. The orthologs of the two target genes are also highly abundant in the adult*N\. americanus*ESP proteome[26](https://www.nature.com/articles/s41467-026-73447-9#ref-CR26), suggesting utility across hookworm species\. The orthologous GSH2 sequence from the*N\. americanus*genome assembly \(PRJNA72135\.WBPS18\) is provided in Supplementary Note[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM1), and all BLAST results for all genes are provided in Supplementary Data[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\.
For both GSH1 and GSH2, CRISPR/Cas9 chromosomal cleavage was compared using combinations of two and three overlapping gRNAs, with ribonuclear protein\-sgRNA complexes \(RNPs\) delivered by electroporation \(Bio\-Rad Gene Pulser\) or by lipofection\. The gRNAs were identified using CHOPCHOP[27](https://www.nature.com/articles/s41467-026-73447-9#ref-CR27); two or more overlapping gRNAs were used, based on previous success with overlapping gRNAs performing better compared to a single gRNA in other helminth species, including*Schistosoma mansoni*[28](https://www.nature.com/articles/s41467-026-73447-9#ref-CR28),[29](https://www.nature.com/articles/s41467-026-73447-9#ref-CR29)\. Sequences of the four sets of overlapping gRNAs are provided in Supplementary Note[2](https://www.nature.com/articles/s41467-026-73447-9#MOESM1), and their positions in the GSH sequences are detailed in Supplementary Note[3](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\. A targeted amplicon sequencing approach was used to quantify programmed CRISPR/Cas9 chromosomal cleavage, i\.e\., sequence modification events at the predicted cleavage position \(primer sequences provided in Supplementary Note[4](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. We estimated the editing efficiencies by comparing the rate of deletions between the treatment and the control group\. Following electroporation of the 12,934,578 hookworm\-derived reads from GSH1, 0\.04332% showed GSH1 deletions, compared to 0\.00371% for control hookworms, representing an 11\.7\-fold increase in deletion rate\. Likewise, for GSH2, of the 20,913,410 reads sequenced from electroporated hookworms, 0\.09598% showed GSH2 deletions, compared to 0\.00683% of the 6,487,031 reads from control, representing a 14\.1\-fold increase in the deletion rate compared to control and a 2\.2\-fold increase compared to GSH1\. The distribution of deletion lengths and sequence alignments showed one consistent 56 bp deletion spanning 34 bp upstream and 22 bp of the GSH1 cleavage site, flanked by nucleotides ATT \(Supplementary Fig\.[3a](https://www.nature.com/articles/s41467-026-73447-9#MOESM1); Supplementary Note[5a](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\), indicative of possible CRISPR\-induced microhomology\-mediated end joining \(MMEJ\)[30](https://www.nature.com/articles/s41467-026-73447-9#ref-CR30)\. In contrast, targeting of GSH2 induced a range of small deletions ranging from 1–13 bp as expected following programmed CRISPR cleavage and resolution of the chromosomal lesion by non\-homologous end joining\. Notably, a consistent 35 bp deletion represented ~15% of all indels observed that spanned the programmed cleavage site \(Supplementary Fig\.[3b](https://www.nature.com/articles/s41467-026-73447-9#MOESM1); Supplementary Note[5b](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\.
Accordingly, in the downstream investigation, we targeted GSH2 \(rather than GSH1\) with our RNPs, based on the increased number of deletions and the conformity of CRISPR indel size distribution\. To target GSH2 in*A\. ceylanicum*adults, we used lipofection\-based transfection based on a protocol for lipofection of larval*Brugia malayi*\(lymphatic filarial worm[31](https://www.nature.com/articles/s41467-026-73447-9#ref-CR31); see Methods\)\. Lipofection encapsulates the RNP complex, so we considered that it may protect the RNP complex from secreted hydrolases, while providing all the CRISPR components together to the localized cells\. However, of the 7,504,603 reads sequenced from the transfected adult*A\. ceylanicum*, only 0\.00556% showed GSH2 deletions, which was not an improvement compared to the control\.
Collectively, these data validated that \(1\) the GSH2 locus as a highly accessible region for CRISPR\-mediated editing in*A\. ceylanicum*, and \(2\) electroporation outperformed lipofection in the delivery of RNP mediated CRISPR gene editing at GSH2\.
### Recombinant s16\-HuScFv mixed with hookworm ESPs neutralizes TTX
Hookworms evolved a sophisticated secretory system that delivers \>800 proteins[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\. To assess feasibility of the approach, we first evaluated potentially negative impacts of native ESP, which includes numerous proteases[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17), on recombinant anti\-tetrodotoxin human single chain antibody fragment \(s16\-HuScFv[13](https://www.nature.com/articles/s41467-026-73447-9#ref-CR13); purity of 95% by SDS\-page; Supplementary Fig\.[4](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. This assessment used a nerve cell based assay where osmotic cell lysis is used to quantify TTX binding to ion channels[13](https://www.nature.com/articles/s41467-026-73447-9#ref-CR13)\(see Methods and schematic for all testing in Supplementary Fig\.[5](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. As shown in Fig\.[2](https://www.nature.com/articles/s41467-026-73447-9#Fig2), the recombinant s16\-HuScFv, when incubated with 200 nM TTX before in vitro exposure to Neura\-2a cells, significantly neutralizes TTX in a dose\-dependent manner \(*P*= 1\.0 × 10\-3for neutralization difference between 700 nM and 3500 nM of s16\-HuScFv; ANOVA with Tukey HSD post\-hoc comparisons\)\. The presence of 0\.5 µg/ml hookworm ESPs \(see Methods\) at both s16\-HuScFv concentrations did not significantly reduce s16\-HuScFv neutralization activity at either concentration \(*P*= 0\.90 at 700 nM and*P*= 1\.0 at 3500 nM\), and the dose\-dependence of s16\-HuScFv activity was maintained in the presence of the hookworm ESPs \(*P*= 3\.2 × 10\-4\)\. Additionally, fluorescence signals did not change after as little as 6 hours, up to 72 hours \(Supplementary Fig\.[6](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. Overall, these results demonstrated that the presence of hookworm ESPs did not interfere with the ability of s16\-HuScFv to neutralize TTX in a dose\-dependent manner, even over an extended period\.
**Fig\. 2: In vitro neutralization of TTX by recombinant s16\-HuScFv antibody fragments in the presence of hookworm excreted/secreted products \(ESPs\)\.**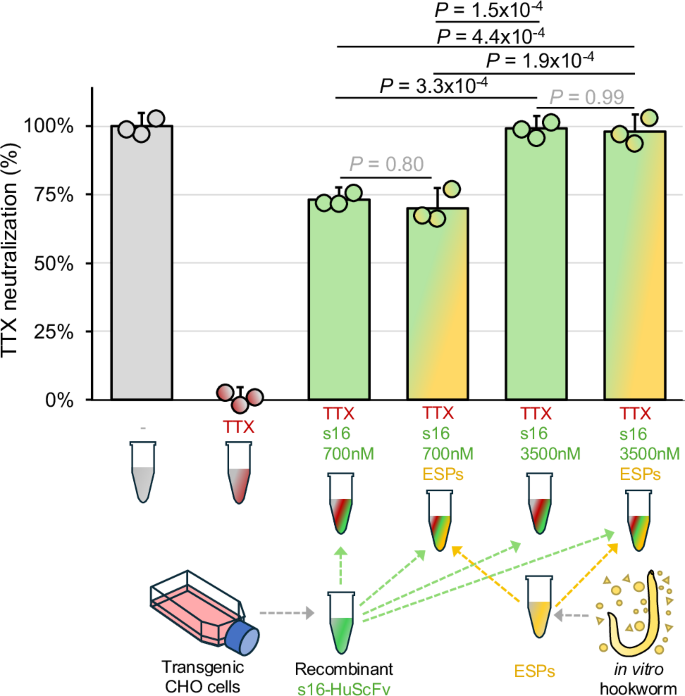The alternative text for this image may have been generated using AI\.
[Full size image](https://www.nature.com/articles/s41467-026-73447-9/figures/2)
TTX neutralization was measured by osmotic lysis of Neura\-2a cells by the ion\-channel mediators veratridine \(V; 0\.2 mM\) and ouabain \(O; 0\.4 mM\) using the Promega CellTox Green Cytotoxicity Assay for up to 72 hours\. TTX neutralization is quantified between 0% \(V \+ O \+ TTX, with no ScFv, red\) and 100% \(V \+ O, with no TTX, grey\) based on relative fluorescence units, adjusted for dilution factors\. Bars represent the standard deviation of the mean based on three replicates per sample\. Samples included s16\-HuScFv at 700 nM and 3,500 nM \(green\), with and without 0\.5 µg/mL hookworm excretory/secretory proteins \(ESPs; yellow\) in the reaction\. A one\-way ANOVA was performed to compare the s16\-HuScFv groups \(*n*= 3 per group, degrees of freedom=3 between and 6 within;*F*\-statistic=17\.5,*P*= 2\.7 × 10\-5, effect size=0\.766, Cohen’s*f*= 1\.81\), with Tukey’s HSD applied for post\-hoc comparisons\. The two technical control samples defining 0% and 100% neutralization for the assay were excluded from the ANOVA\. Source data are provided as a Source Data file\.
### Engineering and delivery of transgene construct encoding s16\-HuScFv
Our goal was to bioengineer a hookworm to produce and secrete a model antibody, anti\-tetrodotoxin human single\-chain variable fragment antibody s16\-HuScFv[13](https://www.nature.com/articles/s41467-026-73447-9#ref-CR13)\. The*s16\-HuScFv*transgene had VH and VL sequences linked into DNA sequences coding for human single\-chain variable fragments \(*HuScFv*\) using a polynucleotide linker \(G4S\)3 and a C\-terminal hexa\-histidine motif \(Fig\.[1](https://www.nature.com/articles/s41467-026-73447-9#Fig1)a,[b](https://www.nature.com/articles/s41467-026-73447-9#Fig1)\)\. The assembly of the double\-stranded donor construct was based on reports of successful use of individual components in previous studies and on new empirical data\. To ensure s16\-HuScFv secretion from the hookworm to its environment, we identified signal peptides for secretion to be tested, based on seven prioritized genes \(Supplementary Fig\.[7a](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\): \(i\)*Ancylostoma caninum*ASP\-2, a highly secreted protein from hookworms*A\. caninum*[32](https://www.nature.com/articles/s41467-026-73447-9#ref-CR32)and*N\. americanus*[33](https://www.nature.com/articles/s41467-026-73447-9#ref-CR33)in the literature, \(ii\) proteins with signal peptides that were among the top 25% most abundant ESPs in adult*A\. ceylanicum*[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17), the top 10% most highly expressed across the*A\. ceylanicum*life cycle[14](https://www.nature.com/articles/s41467-026-73447-9#ref-CR14), and those without detectable expression in the intestine of adult male*A\. ceylanicum*[34](https://www.nature.com/articles/s41467-026-73447-9#ref-CR34)\. This last criterion ensured that secretion is not primarily into the worm’s intestine, where digestion of the host blood meal occurs, but rather directly outside of the worm into the host milieu\. The five signal peptide sequences that were selected for experimental evaluation included: An ASP\-1 ortholog \(ACEY\_09708\-1\), and AP\-1 \(ACEY\_09599\-1\), an anti\-coagulant protein that targets host coagulation factors[35](https://www.nature.com/articles/s41467-026-73447-9#ref-CR35); and \(iii\) the signal peptides from the proteins corresponding to the two GSH target genes from the analysis presented above\. Experimental validation was performed by western blot detection of transgenic GFP protein in HEK 293 T cell supernatant and adult hookworm ESPs \(Supplementary Fig\.[7b](https://www.nature.com/articles/s41467-026-73447-9#MOESM1);*see*Methods\)\. The ASP\-1 signal peptide resulted in the highest transgenic protein detection in both HEK 293 T cell supernatant and in ESP collected from electroporated adult*A\. ceylanicum*worms\. Based on these results, the ASP\-1 signal peptide sequence was used to direct the antibody into the hookworm secretory pathway\. Furthermore, to facilitate optimal protein synthesis of the transgene\-encoded antibody, we modified the translation initiation site by insertion of two extra residues \(Lys\-Ile\) at amino positions 3 and 4 of the signal peptide sequence \(MVKITYNIAFFVLLAASSSVVT\), a modification demonstrated to improve protein synthesis[36](https://www.nature.com/articles/s41467-026-73447-9#ref-CR36)\. This transgene expression cassette \(NH2\-ASP1 Signal\-s16VH\-\(G4S\)3 Linker\-s16VL\-HisTag\-COOH; Fig\.[1b](https://www.nature.com/articles/s41467-026-73447-9#Fig1)\) was included in the configurable chassis for CRISPR/Cas catalyzed insertion into the germline of*A\. ceylanicum*\.
The synthetic plasmid transgene construct also included the commonly\-used human cytomegalovirus \(CMV\) promoter sequence that has been shown to produce robust gene expression and protein production in other helminths, including the model nematode organism*C\. elegans*[37](https://www.nature.com/articles/s41467-026-73447-9#ref-CR37), the tapeworm*Taenia crassiceps*[38](https://www.nature.com/articles/s41467-026-73447-9#ref-CR38)and the blood fluke*S\. mansoni*[39](https://www.nature.com/articles/s41467-026-73447-9#ref-CR39)\. In the absence of a well\-annotated and experimentally validated endogenous*A\. ceylanicum*promoter, we placed the CMV promoter sequence before the*s16\-HuScFv*to drive the transgene expression\. Likewise, the commonly used bGH/poly\(A\)[40](https://www.nature.com/articles/s41467-026-73447-9#ref-CR40)terminator sequence was also placed after the transgene to terminate transcription\. Finally, the construct also included 600 bp left homology and right homology arms \(HAs\) that are specific for homology\-directed repair following programmed CRISPR cleavage at GSH2\. Complete*A\. ceylanicum*GSH2 genomic sequences, including indications of HA and guide RNA sequences, are provided in Supplementary Note[3](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\. The transgene chassis \(i\.e\., donor DNA template, Fig\.[1b](https://www.nature.com/articles/s41467-026-73447-9#Fig1)\) was delivered as linearized, double\-stranded DNA, 3\.2 kb in length \(Supplementary Fig\.[8](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. Linearized DNA \(as opposed to circular plasmid DNA\) and long 600 bp HAs were used based on our CRISPR\-based transgene KI in*S\. mansoni*[28](https://www.nature.com/articles/s41467-026-73447-9#ref-CR28),[29](https://www.nature.com/articles/s41467-026-73447-9#ref-CR29)\.
Since transgenesis has yet to be reported in hookworms[12](https://www.nature.com/articles/s41467-026-73447-9#ref-CR12), to identify optimal electroporation\-based transgene delivery to the genome of*A\. ceylanicum*eggs, we compared the performance of 24 different preprogrammed protocols of the Neon electroporation system[41](https://www.nature.com/articles/s41467-026-73447-9#ref-CR41)\. The evaluated electroporation conditions for the integration of DNA into*in vitro\-*laid*A\. ceylanicum*eggs included varying pulse number, pulse voltage, and pulse width \(time in milliseconds\)[41](https://www.nature.com/articles/s41467-026-73447-9#ref-CR41)\. Transgene integration was confirmed by PCR using a transgenic forward primer and downstream genomic reverse primer, and yielded amplicons of the expected size of ~350 bp for conditions 12, 13, 14, 17 and 18 \(Supplementary Fig\.[9a](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. These results were confirmed with a duplicate round of validation for conditions 12\-14 and 17\-18 \(Supplementary Fig\.[9b](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\), after which conditions 12 and 13 were further validated by PCR using two different sets of primers \(Supplementary Fig\.[9c](https://www.nature.com/articles/s41467-026-73447-9#MOESM1); Supplementary Note[6](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\), resulting in the amplicons of the expected size targets of 850 and 550 bp for protocol 13 \(*see*Methods\) Accordingly, we deployed protocol 13 for transgene delivery in downstream studies\.
Our findings demonstrated that successful and robust transgene delivery in immature eggs of*A\. ceylanicum*was achieved with electroporation and Neon protocol 13 \(1100 volts, 20 milliseconds, 2 pulses\); confirmed through PCR validation and repeatable amplification of target sequences\. While we used in vitro laid eggs from adult female worms in culture medium for identification of the Neon electroporation protocol 13 for delivery of the transgene construct, these immature eggs are collected in low numbers and are more fragile and less likely to develop into infective larvae\. Therefore, mature eggs recovered from hamster feces were used for all other subsequent experiments, as many survive and develop rapidly into infective L3s\. For visualization of*A\. ceylanicum*egg development progression, we isolated immature eggs from the hamster intestine to observe the early stages of embryogenesis \(Supplementary Fig\.[10a](https://www.nature.com/articles/s41467-026-73447-9#MOESM1),[b](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\), relative to the mature eggs collected from the feces \(Supplementary Fig\.[10c](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)and[d](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\.
### Transgenesis and vertical transmission of s16\-HuScFv in hookworms
F0 wild\-type mature eggs were transfected \(Fig\.[3a](https://www.nature.com/articles/s41467-026-73447-9#Fig3)\) using the Neon electroporation protocol 13 with Cas9/RNP1/RNP2/s16\-HuScFv construct donor DNA deployed with two overlapping gRNAs targeting GSH2 for programmed CRISPR/Cas knock\-in\. Each egg pool \(three distinct biological replicates\) was divided into two groups for subsequent analysis; genomic DNA \(gDNA\) was extracted from one group of ~50 pooled eggs and evaluated for the presence of transgene, and a second group of ~200 eggs was cultured on nematode growth medium \(NGM\) plates to obtain the infective third\-stage larvae \(iL3\)\. Three hamsters were infected with these iL3 with the aim of establishing adult transgenic parental F0 hookworms and sexual reproduction to release F1 generation eggs\. Feces from these hamsters were sampled on 18,19 and 20 days post infection \(dpi\) to recover putatively transgenic F1 eggs\. The WT and transgenic F1 eggs were split into two groups: one evaluated for the presence of the*s16\-HuScFv*transgene following targeted KI into the genome, and the second to evaluate their subsequent progression through early larval development\. Following euthanasia, these hamsters were necropsied, the resulting F0 adult hookworms were retrieved from the small intestine, and genomic DNA from these adult hookworms were evaluated for the transgene integration at GSH2\.
**Fig\. 3: Targeted knock\-in of s16\-HuScFv in hookworm eggs and evidence for germ line transgenesis\.**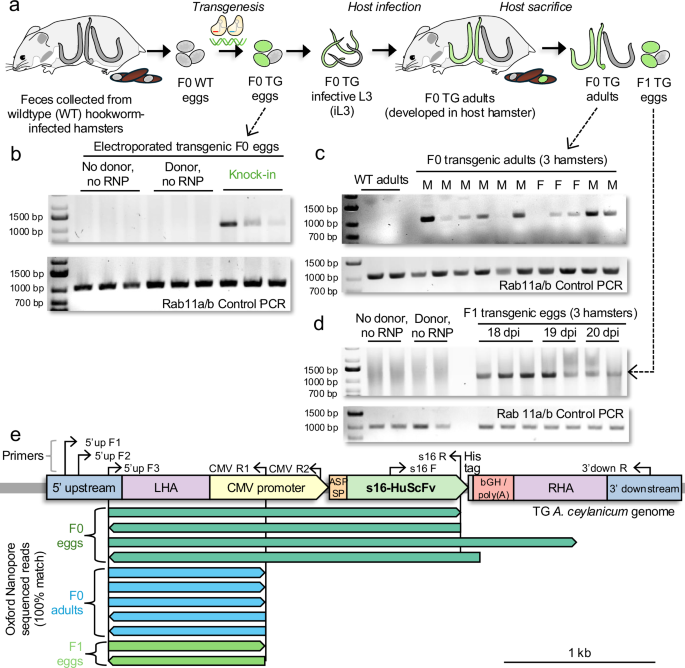The alternative text for this image may have been generated using AI\.
[Full size image](https://www.nature.com/articles/s41467-026-73447-9/figures/3)
**a**Schematic of*A\. ceylanicum*programmed CRISPR\-Cas knock\-in \(KI\) approach using the hamster infection model\.**b**Targeted KI of transgene confirmed in F0 eggs \(*n*= 3\) by PCR using primers 5’up F3 & CMV R1 \(expected 1061 bp amplicon\)\. Controls included electroporation without donor DNA or ribonucleoprotein \(RNP\) complex \(*n*= 3\), and electroporation with donor DNA but without RNP complex \(*n*= 3\)\. The integrity of the DNA was confirmed by PCR amplification of the*Rab11a*/*b*constitutive gene \(“Control PCR”, expected 1000 bp amplicon\)\.**c**Targeted KI of the transgene confirmed in F0 adults recovered from intestines of hamsters \(*n*= 3\) by genomic DNA PCR, using 5’up F3 and CMV R1 primers \(expected 1061 bp amplicon\)\. The integrity of the DNA was confirmed by PCR amplification of the*Rab11a*/*b*constitutive gene \(“Control PCR”, expected 1000 bp amplicon\)\.**d**Targeted transgene KI confirmed in F1 eggs from 3 hamster hosts by PCR \(eggs collected on day 18, 19, and 20 of infection\) using 5’up F3 and CMV R1 primers \(expected 1,061 bp amplicon\)\. The integrity of the DNA was confirmed by PCR amplification of the*Rab11/b*constitutive gene \(“Control PCR”, expected 350 bp amplicon\)\.**e**Verification of transgene knock\-in by sequencing of the targeted genomic region using ONT sequencing\. Sequencing was performed on F0 transgenic eggs \(pool of eggs, dark green; 5’up F3 and s16 R primers; \(**b**\)\); F0 adults \(single worms, blue; 5’up F3 and CMV R1 primers; \(**c**\)\) and F1 transgenic eggs \(pool of eggs, light green; 5’up F3 and CMV R1 primers \(**d**\)\)\. The transgene construct is shown in the*A\. ceylanicum*genome, with additional primers used for testing also shown\. All ONT sequencing reads exactly matched the expected transgenic genomic sequences\. Source data are provided as a Source Data file\.
The programmed CRISPR\-directed KI \(i\.e\., the*s16\-HuScFv*transgene integration into the GSH2 genomic locus; Fig\.[3b–d](https://www.nature.com/articles/s41467-026-73447-9#Fig3)\), was assessed by PCR primers designed to flank the junction between the endogenous genomic DNA and the donor sequence \(schematic in Fig\.[3e](https://www.nature.com/articles/s41467-026-73447-9#Fig3)\)\. Forward primers were annealing to the genomic region upstream of the 5’ homology arm \(LHA\), while reverse primers targeted either the CMV promoter or a specific s16\-HuScFv sequence within the donor construct\. These primer pairs were intended to amplify the segment spanning the genomic donor junction, thereby confirming successful transgene integration\. The resulting PCR amplicons were subsequently analyzed by Oxford Nanopore Technology \(ONT\) sequencing to further validate precise insertion of the s16\-HuScFv transgene at the targeted genomic site\. All primer sequences are provided in Supplementary Note[6](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\. We used a forward primer specific for the genomic DNA upstream of the 5’ homology arm \(5’up F3\), paired with a reverse primer specific for the CMV promoter sequence \(CMV R1\) or s16\-HuScFv transgene sequences \(s16 R\)\. In the F0 transgenic eggs, PCR amplicons of the expected sizes of 1061, respectively, were observed \(Fig\.[3b](https://www.nature.com/articles/s41467-026-73447-9#Fig3); upper and middle panel\)\. This band was not observed in the control groups, which included mock treatment \(electroporated eggs without donor and RNPs complexes\) and donor only \(electroporated eggs with donor DNA but without RNPs complexes\) groups, while amplicons specific for the constitutive*Rab11a/b*gene were observed in all samples \(Fig\.[3b](https://www.nature.com/articles/s41467-026-73447-9#Fig3), lower panel\)\.
To evaluate the vertical transmission of*s16\-HuScFv*in the hookworm, transgenesis of*s16\-HuScFv*into the genome of 9 of the 11 F0 adults was confirmed by PCR using 5’ upstream F3 and CMV R1 primers, yielding an amplicon 1,061 bp in length\. The gDNA integrity was validated by PCR targeting the*Rab11a*/*b*constitutive gene \(expected size, 1000 bp; Fig\.[3c](https://www.nature.com/articles/s41467-026-73447-9#Fig3)\)\. Likewise, vertical transgene transmission was validated by obtaining PCR amplicons of the expected size \(1061 bp\) from F1 eggs pooled from three hamsters at 18, 19, and 20 days post\-infection \(Fig\.[3d](https://www.nature.com/articles/s41467-026-73447-9#Fig3); control confirmation using the*Rab11a/b*constitutive gene, expected size, 350 bp\)\. The F1 eggs hatched and developed to the infective L3 stage\. For each of the three sample groups, visible PCR bands at the expected sizes were eluted from the gels and were sequenced by ONT sequencing, to provide validation for successful transgene KI at the GSH2 locus\. Each of the sequenced reads spanning the 5’ upstream genomic sequence and into either the CMV promoter or the*s16\-HuScFv*gene sequence exactly matched the expected sequence at every base position \(Fig\.[3e](https://www.nature.com/articles/s41467-026-73447-9#Fig3); Supplementary Note[7](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\), validating the PCR results indicating both targeted transgenesis and vertical transmission of the transgene\.
The results confirmed the successful integration of the*s16\-HuScFv*transgene into the hookworm genome at the GSH2 target site\. Through PCR amplification, transgenesis was confirmed in F0 eggs, both the adult male and female F0, and F1 generation of eggs, with amplicons of the expected sizes consistently observed in transgenic \(TG\) samples but absent in controls\. The ONT sequencing results confirmed precise transgene insertion at the targeted genomic locus, confirmed by sequence alignment with the expected donor DNA sequences \(Supplementary Note[7d](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. To reiterate, transgenesis and vertical transmission of the transgene were established, as shown by the presence of the*s16\-HuScFv*gene in both F0 adults and F1 eggs, respectively, collected from infected hamsters\.
### s16\-HuScFv expression did not disrupt surrounding gene expression
Given that hookworm transgenesis had not been reported, we proceeded to thoroughly evaluate the effects of transgene KI at GSH2\. To quantify the transcription of*s16\-HuScFv*and evaluate its targeted KI effect on the expression of the surrounding genes, four individual experiments \(four biological replicates\) were performed\. Approximately 20,000 mature eggs per experiment were electroporated with the s16\-HuScFv\-encoding construct, along with four pools of eggs that underwent electroporation without any donor DNA \(mock transgenesis; wild type, WT\)\. PCR confirmed construct insertion in the cohort of transgenic eggs with two discrete primer sets, and without detection in WT;*Rab11a/b*constitutive gene detection confirmed intact*A\. ceylanicum*DNA in all samples \(Fig\.[4a](https://www.nature.com/articles/s41467-026-73447-9#Fig4)\)\. qRT\-PCR analysis using three of the biological replicates with the best QC values each confirmed gene expression of*s16\-HuScFv*in the transgenic eggs, with no detection in the WT samples, relative to the reference gene,*Rab11A*\. The expression levels of surrounding genes,*ACEY\_002221*\(upstream of the GSH insertion site\) and*ACEY\_002225*\(downstream\), were unaffected \(*P*= 0\.40 and 0\.43, respectively\) in the TG eggs compared to the WT eggs, as well as that of a second reference gene,*benA*\(*P*= 0\.40; Fig\.[4b](https://www.nature.com/articles/s41467-026-73447-9#Fig4)\)\. qRT\-PCR was also performed with the same approach on two pools of four adult worms each, which confirmed detection of*s16\-HuScFv*in TG adults but not WT adult worms \(at a lower expression level than in eggs\), and without significant impact on expression levels of neighboring genes or*benA*\(Supplementary Fig\.[11](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. The PCR and qRT\-PCR primer sequences are listed in Supplementary Note[6](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\.
**Fig\. 4: Confirmation of s16\-HuScFv gene expression in transgenic A\. ceylanicum eggs\.**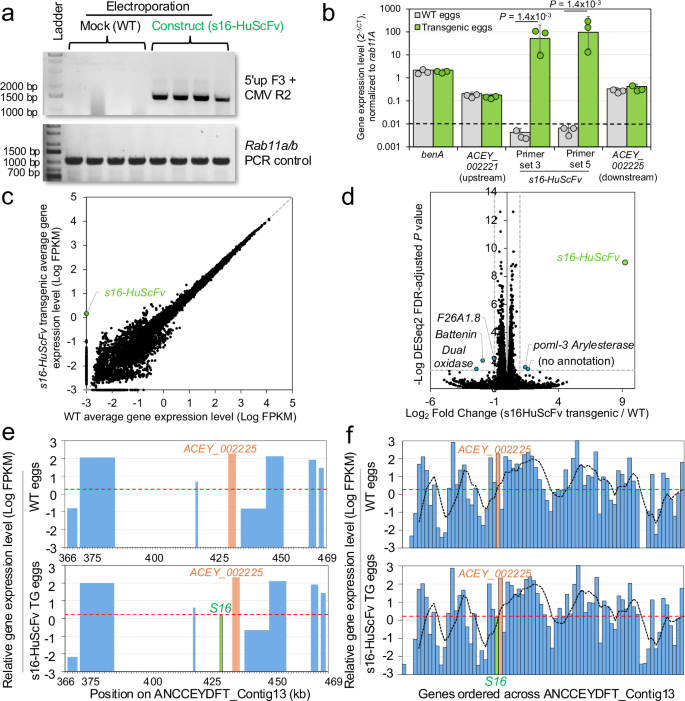The alternative text for this image may have been generated using AI\.
[Full size image](https://www.nature.com/articles/s41467-026-73447-9/figures/4)
**a**Targeted knock\-in of transgene confirmed in F0 eggs by PCR using 5’up F3 & CMV R2 \(expected 1,445 bp amplicon\)\. Controls included mock electroporation without donor DNA or RNP complexes \(WT\)\. The integrity of the DNA was confirmed by PCR amplification of the*Rab11a/b*constitutive gene \(expected 350 bp amplicon\)\. Three samples from each group were selected for qPCR and RNA\-seq validation\.**b**qRT\-PCR\-based confirmation of*s16\-HuScFv*gene expression in transgenic*A\. ceylanicum*eggs, using two sets of qRT\-PCR primers\. Normalization was performed relative to the*R**ab11A*constitutive gene\.*P*values represent two\-sided*t*\-tests with unequal variance \(with FDR correction applied for multiple tests\) using untransformed ∆CT values, the dashed line indicating the qRT\-PCR detection limit threshold, and error bars represent standard deviation of the mean;*n*= 3 per group\. Significant differences were identified for s16\-HuScFv primer set 3 \(FDR\-adjusted*P*= 1\.4 × 10\-3*, t*= 11\.6, effect size = 9\.5\) and set 5 \(FDR\-adjusted*P*= 1\.4 × 10\-3,*t*= 10\.0, effect size = 8\.15\); 95% t confidence interval = \-2\.78:2\.78 and effect size = 4 for both\. Relative expression values in WT and transgenic*s16\-HuScFv*sample sets are also shown for the gene immediately downstream and the upstream gene from the GSH insertion \(primers for the immediately upstream*ACEY\_002197\-1*did not amplify\), to demonstrate a lack of disruption of normal gene expression, and constitutive gene benA expression is shown as an additional validation\.**c**RNA\-seq\-based average relative gene expression levels \(log FPKM\) for all 17,333*A\. ceylanicum*genes in the three WT samples and the s16\-HuScFv transgenic samples\. ND=Not detected\.**d**Volcano plot depicting log2fold change values and \-log values of FDR\-corrected*P*values for significant differential gene expression according to DESeq2\. Dashed lines indicate thresholds for significance \( \| Fold change \| ≥ 1,*P*≤ 0\.05\)\. Genes exceeding significance thresholds are shown with blue dots, and s16\-HuScFv, only detected in transgenic eggs, is shown with a green dot\.**e**Relative gene expression levels of genes surrounding the GSH region, in WT eggs and s16\-HuScFv transgenic eggs\. X\-axis positions indicate genomic positions on the genomic contig containing the GSH region \(*ANCCEYDFT\_Contig13*\)\. The dashed red line indicates the average expression level of all genes\.**f**Relative gene expression levels of all genes on*ANCCEYDFT\_Contig13*\. The red dashed line indicates the average expression level of all genes, and the black dashed line indicates the running average of five genes across the contig\. Orange bars indicate ACEY\_002225 target gene, green bars indicate s16\-HuScFv\. Source data are provided as a Source Data file\.
Next, bulk RNA\-seq analysis of gene expression was performed for WT and TG hookworm \(*n*= 3 biological replicates per group from each of the qRT\-PCR sample sets;*n*= 6 total\), generating an average of 60\.3 million read pairs per sample with detection of an average of 84\.4% of all 18,776 protein\-coding genes in the hookworm genome\. Gene expression levels for all genes correlated very strongly between the WT and TG sample groups \(Pearson correlation=0\.998 on natural scale, and 0\.985 on log scale; Fig\.[4c](https://www.nature.com/articles/s41467-026-73447-9#Fig4)\)\. Differential gene expression analysis with DESeq2[42](https://www.nature.com/articles/s41467-026-73447-9#ref-CR42)identified only six genes with significantly different expression levels between the WT and TG samples \(out of 16,066 total detected genes; minimum 2\-fold expression difference and FDR\-adjusted*P*value ≤ 0\.05\), including*s16\-HuScFv*\(no detection in WT, expressed higher than 41% of other genes in the three TG biological replicates;*P*= 1\.0 × 10−9\) and five other genes with low average expression values \(all expressed lower than at least 60% of all genes; Fig\.[4d](https://www.nature.com/articles/s41467-026-73447-9#Fig4)\), all of which are on genomic contigs separate from that of GSH2 insertion region and do not appear to be critical for survival based on deduced functional annotations\. These include dual*oxidase*/*bli\-3*, which generates reactive oxygen species as an innate immune mechanism[43](https://www.nature.com/articles/s41467-026-73447-9#ref-CR43),*battenin/cln\-3\.2*\(involved in aging[44](https://www.nature.com/articles/s41467-026-73447-9#ref-CR44)\),*arylesterase/poml\-3*\(functions unknown in nematodes\), and two genes without functional annotation \(an ortholog of F26A1\.8 and an*A\. ceylanicum*\-specific protein\)\. No significant changes in expression of genes immediately surrounding the GSH2 region \(Fig\.[4e](https://www.nature.com/articles/s41467-026-73447-9#Fig4)\) or across the entire contig containing the GSH \(Fig\.[4f](https://www.nature.com/articles/s41467-026-73447-9#Fig4)\) were observed, with expression patterns remaining consistent across the groups\. Supplementary Data[4](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)includes the read count metadata and NCBI SRA accessions \(BioProject PRJNA1328670\) while Supplementary Data[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)lists all read counts, normalized expression values and differential expression statistics\.
Together, the gene expression analysis results confirmed relatively high gene expression level of the*s16\-HuScFv*in the TG samples, and absence from WT samples, without impact on expression of the surrounding genes, and therefore no expected changes to worm fitness as a result of KI of the transgene into the GSH2 target site\.
### TTX neutralization by hookworm\-produced s16\-HuScFv in hamster serum
We assessed production, secretion and activity of s16\-HuScFv by transgenic hookworm into the venous bloodstream by collecting serum from hamsters infected with L3s developed from electroporated TG and WT eggs, at 22 days post\-infection \(*n*= 4 biological replicates per group; see Methods\)\. First, mass spectrometry proteomics using the Spectronaut[45](https://www.nature.com/articles/s41467-026-73447-9#ref-CR45)directDIA approach was utilized to detect serum protein abundance, including s16\-HuScFv and a background of all*A\. ceylanicum*proteins and all host hamster proteins\. This approach detects peptides in every sample at each mass range, resulting in very high sensitivity, and some expected background detection noise[46](https://www.nature.com/articles/s41467-026-73447-9#ref-CR46)\. Two of the four s16\-HuScFv transgenic hookworm\-infected replicates had substantially higher detection of s16\-HuScFv than the WT background controls \(3\.3\-fold and 6\.4\-fold higher detection\)\. Differential expression using limma[47](https://www.nature.com/articles/s41467-026-73447-9#ref-CR47)using these two samples and the four WT controls yielded a 4\.71\-fold increase in s16\-HuScFv in the transgenic samples \(*P*= 2\.6 × 10−3\), with none of the other 27 detected*A\. ceylanicum*proteins being identified as significantly more abundant in the transgenic samples\. Two of these 27 proteins were previously detected in adult ESPs using mass spectrometry[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17), and 7 of the 27 were orthologous to*Haemonchus contortus*extracellular vesicle \(EV\) proteins[48](https://www.nature.com/articles/s41467-026-73447-9#ref-CR48)\(26\.9%, vs 4\.4% expected by random chance;*P*= 1\.1 × 10−5for enrichment, binomial distribution test\)\. This included fructose\-bisphosphate aldolase \(ACEY\_14462\-1\), which was also an ortholog of one of the 81 EV proteins detected in the rodent hookworm*Nippostrongylus brasiliensis*[49](https://www.nature.com/articles/s41467-026-73447-9#ref-CR49)\. This protein and heat shock protein 90 \(ACEY\_06074\-1\) were also among five EV proteins conserved across cestodes, nematodes, and trematodes identified in an evolutionary conservation analysis[50](https://www.nature.com/articles/s41467-026-73447-9#ref-CR50)\. These results suggest that*A\. ceylanicum*EV protein promoters, signal peptides for secretion, and GSH regions may be useful for optimizing secretion into the host\. Detection level details and a complete list of all hookworm and host proteins detected in each sample are provided in Supplementary Data[5](https://www.nature.com/articles/s41467-026-73447-9#MOESM3), and raw proteomics data are accessible on iProX \(accession IPX0014753001\)\.
Next, we measured TTX neutralization by hookworm\-produced s16\-HuScFv circulating in hamster blood using the neuronal cell osmotic cell lysis assay \(schematics shown in Supplementary Fig\.[5](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. Sera collected from hamsters infected with wildtype iL3 or s16\-HuScFv transgenic iL3 at 22 dpi \(Fig\.[5a](https://www.nature.com/articles/s41467-026-73447-9#Fig5)\) were evaluated, and showed that sera from hamsters infected with transgenic iL3 were capable of neutralizing approximately 16\.3% of TTX \(*P*= 9\.1 × 10−5, FDR\-corrected one\-tailed*t*\-test; Fig\.[5b](https://www.nature.com/articles/s41467-026-73447-9#Fig5);*t*=−10\.4909, 95% confidence interval = \-2\.015:∞, effect size=8\.01,*n*= 3 controls and*n*= 4 TG samples, degrees of freedom = 5\), while sera from wildtype hookworm controls showed zero neutralization \(*P*= 0\.5, FDR\-corrected one\-tailed*t*test,*t*= 6\.6 × 10\-17, 95% confidence interval = −2\.1318:∞, effect size=5\.4 × 10−17,*n*= 3 per sample group, degrees of freedom = 4\)\. ELISA\-based quantification of s16\-HuScFv was attempted for WT and transgenic adult*A\. ceylanicum*ESP and for sera from hamsters infected with both groups of worms, using anti\-HIS capture antibody and ab\-Goat Anti\-Human IgG detection antibody \(F\(ab\)2\-fragment specific;*see*Methods for complete details\)\. Based on analysis of the standard curve using recombinant s16\-HuScFv, the ELISA had a limit of detection of ~79\.2 nM \(using the regression\-based approach; LOD = 3\.3 × σ / slope\) and a limit of quantification of ~239\.8 nM \(LOQ = 10 × σ/slope\)\. Our results suggest that the s16\-HuScFv concentration in the hamster serum is below the LOD \(Supplementary Fig\.[12](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\); nonetheless, we demonstrated functional human antibody in the circulation of hamsters infected with transgenic L3s based on the results from the TTX neutralization assay\.
**Fig\. 5: Neutralization of TTX by the serum of hamsters infected with transgenic hookworms\.**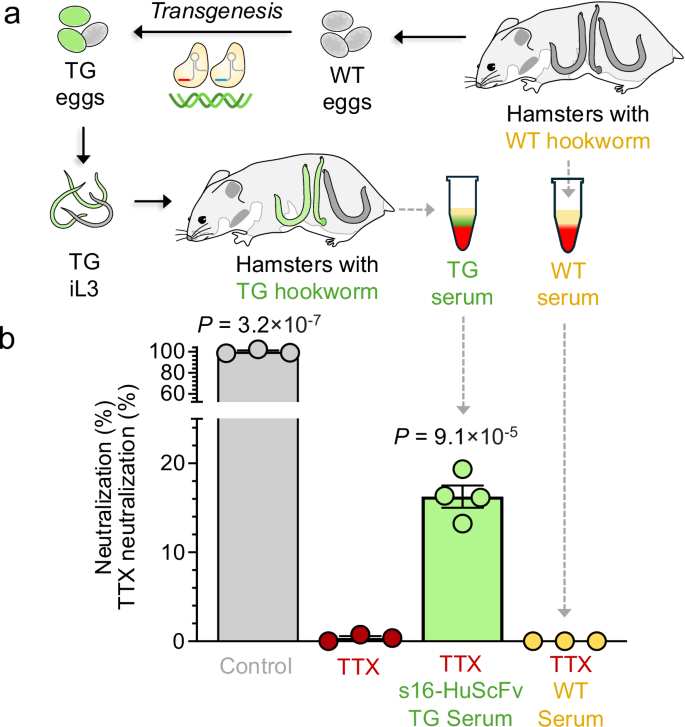The alternative text for this image may have been generated using AI\.
[Full size image](https://www.nature.com/articles/s41467-026-73447-9/figures/5)
**a**Schematic of programmed CRISPR\-Cas knock\-in using a hamster model for human*A\. ceylanicum*infection\. Serum was collected from hamsters infected with WT hookworms and from hamsters with TG hookworms\.**b**Neutralization of TTX by serum from hamsters infected with wildtype and transgenic*A\. ceylanicum*iL3\. TTX neutralization was measured by osmotic lysis of Neura\-2a cells by the ion\-channel mediators veratridine \(V; 0\.2 mM\) and ouabain \(O; 0\.4 mM\) using the Promega CellTox Green Cytotoxicity Assay for up to 72 hours\. TTX neutralization is quantified between 0% \(V \+ O \+ TTX, with no ScFv, red\) and 100% \(V \+ O, with no TTX, grey\) based on relative fluorescence units, adjusted for dilution factors\. Bars represent standard error of the mean \(SEM\) based on three technical replicates for the technical controls, and the standard deviation of the mean for distinct biological replicates for the serum samples from hamsters infected with WT*A\. ceylanicum*\(*n*= 3, yellow\) and electroporated transgenic*A\. ceylanicum*expressing s16\-HuScFv \(*n*= 4, green\)\. FDR\-corrected*P*values are shown for one\-tailed*t*\-tests comparing each experimental group to the V \+ O \+ TTX neutralization controls \(red\)\. Significance was observed for V \+ O control \(FDR\-adjusted*P*= 3\.2 × 10−7*, t*= −85\.3,*t*95% confidence interval = \[−2\.132: ∞\], effect size = 69\.67, degrees of freedom = 5\) and for s16\-HuScFv transgenic serum \(FDR\-adjusted*P*= 9\.1 × 10\-5*, t*= −10\.67,*t*95% confidence interval = \[−2\.015: ∞\], effect size = 8\.15, degrees of freedom = 5\)\. Source data are provided as a Source Data file\.
Our findings confirmed that transgenic hookworms successfully produce and secrete s16\-HuScFv into hamster circulation, resulting in a partial neutralization of TTX, while WT hookworms show no neutralization, thus demonstrating the functional activity of the hookworm\-produced s16\-HuScFv antibody\.
## Discussion
Here, we report the genetic modification of the human hookworm*Ancylostoma ceylanicum*, into a living biofactory capable of producing and delivering a functionally active human single\-chain variable fragment antibody \(s16\-HuScFv\) via bioengineered secretomes\. The antibody, s16\-HuScFv, originated in a human ScFv phage display library, and has been reported to neutralize tetrodotoxin \(TTX\) and to rescue intoxicated mice from TTX\-mediated lethality[13](https://www.nature.com/articles/s41467-026-73447-9#ref-CR13)\. TTX and its potent neurotoxin analogs are weaponizable agents that block voltage\-gated sodium ion channel activity in nerve cells[51](https://www.nature.com/articles/s41467-026-73447-9#ref-CR51), resulting in organ paralysis and death, with no commercial antidote available[52](https://www.nature.com/articles/s41467-026-73447-9#ref-CR52)\. Our goal was to engineer hookworms capable of producing and delivering, into the host circulation, a functionally active human antibody that is able to neutralize TTX\. Given that adult hookworms secrete many hundreds of ESPs[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17), we first demonstrated that s16\-HuScFv is stable and retains neutralization activity in the presence of these ESPs\. This was an essential first step that provided the feasibility of the study\.
While gene\-editing technologies have revolutionized biomedical science, progress in the field of helminthology has been far more restrained and, for human hookworms, essentially nonexistent[12](https://www.nature.com/articles/s41467-026-73447-9#ref-CR12)\. This study identifies and showcases the utility of GSH2 as a safe genomic locus in the*A\. ceylanicum*genome for transgene KI and that electroporation, as a mode of transgene delivery, outperforms lipofection\. The ASP\-1 signal peptide selected for secretion of the transgene resulted in detectable secretion, and the*ASP\-1*gene was more recently found to be one of the 53 single\-cell transcriptomic marker genes for the pharyngeal gland in both adult female and male*A\. ceylanicum*[53](https://www.nature.com/articles/s41467-026-73447-9#ref-CR53), \(17\.3\-fold and 24\.8\-fold higher compared to cells from other tissues, respectively\), increasing the likelihood that most of the s16\-HuScFv protein is directed into a secretory pathway rather than remaining in the worm\. In addition to revealing no detectable effects on surrounding or global gene expression, our experimental results show that iL3 derived from transgenic eggs are fit and able to complete the full life cycle in the hamster, which represents the ultimate assessment of the biological impact of transgene expression on worm fitness\. The motility of F0 transgenic adult worms was comparable to that of wild\-type adult worms\. Transgenic F0 females laid eggs in vitro, and these F1 eggs were confirmed to be transgenic by PCR and ONT sequencing and progressed through development to iL3 larvae, demonstrating vertical transmission in hookworms\.
Our results, when considered in the context of the extensive knowledge of hookworm biology, life cycle, and co\-evolution of these parasites with their mammalian hosts, demonstrate that engineered hookworms can serve as a sustained, slow\-release drug production and delivery platform\. Hookworms have coexisted with mammalian hosts, potentially since vertebrates first colonized the land and became vulnerable to parasitic nematodes 350 million years ago[54](https://www.nature.com/articles/s41467-026-73447-9#ref-CR54), which has enabled their long\-term survival, often for years, within the human host\. Moreover, numerous studies have shown that controlled hookworm infections are safe and well tolerated \(e\.g\., refs\.[4](https://www.nature.com/articles/s41467-026-73447-9?error=cookies_not_supported&code=1270f4db-8d1f-4c44-88bf-78d0133011e7#ref-CR4),[5](https://www.nature.com/articles/s41467-026-73447-9?error=cookies_not_supported&code=1270f4db-8d1f-4c44-88bf-78d0133011e7#ref-CR5),[6](https://www.nature.com/articles/s41467-026-73447-9#ref-CR6)\) and can confer therapeutic benefits, particularly in patients with chronic inflammatory diseases, by restoring homeostasis within the gut microenvironment[5](https://www.nature.com/articles/s41467-026-73447-9#ref-CR5),[55](https://www.nature.com/articles/s41467-026-73447-9#ref-CR55),[56](https://www.nature.com/articles/s41467-026-73447-9#ref-CR56), and without negatively altering the gut microbiota[57](https://www.nature.com/articles/s41467-026-73447-9#ref-CR57)\. Collectively, these results highlight the promise of hookworms as an emerging biofactory for the production and delivery of pharmaceuticals, creating a foundation for the delivery of not only therapeutics but also other protein products, such as desensitizing allergens for food allergy and celiac disease[58](https://www.nature.com/articles/s41467-026-73447-9#ref-CR58)\.
Here we present some level of optimization to increase efficiency of transgenesis as well as optimization of the transgene cassette and the configurable chassis that could result in producing and secreting more therapeutic protein, including selection of GSH, signal peptide for secretion, and modification of the translational site[36](https://www.nature.com/articles/s41467-026-73447-9#ref-CR36)\. In other parasitic nematode species, further codon optimization may be considered, however, codon bias in*A\. ceylanicum*has been shown to closely resemble human codon bias, with a preference for GC\-ending codons, particularly C\-ending codons[59](https://www.nature.com/articles/s41467-026-73447-9#ref-CR59), consistent with the codon usage in this study\. Nevertheless, further improvements could be made by choosing inducible/stronger promoters, more efficient secretion signals, increased serum half\-life of biologics, or through engineered selections for transgenic hookworms to obtain more homogenous populations\. Quantification of the produced secreted therapeutics following these optimizations will be critical for evaluating their efficiency\. Clinically relevant serum concentrations of some therapeutic molecules might be beyond the capability of the helminth when administered at tolerable doses, but the hookworm system may produce substantially higher concentrations of target proteins in the gastrointestinal \(GI\) tract, where the adult hookworms reside, positioning it as an ideal system for targeting GI conditions\. Additionally, as research progresses, it will be essential to ensure that these transgenic organisms do not have unintended ecological or human health impacts, maintaining a balance between innovation and safety\. Hookworms do not reproduce within an infected host, since eggs need to embryonate and hatch outside, and the larvae molt to the L3 stage before becoming infective[60](https://www.nature.com/articles/s41467-026-73447-9#ref-CR60), so once transgenic product secretion rates are carefully quantified in an optimized system, the dosage of product can be controlled and calibrated by limiting the number of larvae used for the infection\. Subsequent infection can be prevented through proper hygiene, and clearance of infection is simple and anodyne using an oral medication \(e\.g\., albendazole 400 mg once[56](https://www.nature.com/articles/s41467-026-73447-9#ref-CR56)\)\. Future studies can also address biocontainment of the genetically modified organism \(GMO\) by engineering suicide genes and/or inducible promoters into the transgene\.
This study demonstrates CRISPR\-assisted establishment of a transgenic hookworm and vertical transmission of the transgene from the parental \(P0/F0\) to the F1 generation\. However, an*A\. ceylanicum*s16\-HuScFv transgenic line warrants detailed multi\-generation monitoring and analysis, which will require substantial research beyond the scope of this report\. Stable vertical transmission of transgenes has been reported in*Strongyloides ratti and S\. stercoralis*[61](https://www.nature.com/articles/s41467-026-73447-9#ref-CR61),[62](https://www.nature.com/articles/s41467-026-73447-9#ref-CR62)\. This is not unexpected, given the difficulty in insertion into the germ line in parasitic nematodes other than the*Strongyloides spp*clade, since*Strongyloides spp*can uniquely complete the life cycle without a host, and they anatomically resemble*C\. elegans*, with intragonadal microinjection facilitating transgene delivery directly to the gametes and germ cells\. By contrast, establishing stable transgenic parental and F1 progeny in hookworms presents a far more formidable challenge\. Once established, further studies can be undertaken to demonstrate host protection against challenge with TTX in vivo\.
Several essential components have converged to support the development of transgenic hookworms as a next\-generation platform for drug production and delivery with practical clinical application\. Building on the foundation that experimental human hookworm infection has been shown to be safe and well tolerated, here we present technological, methodological, and conceptual advances that have enabled the establishment of a genetically modified and tractable model system that can produce and deliver biologics\. These advances include improved prediction and validation of target insertion sites, the use of overlapping gRNAs targeting the GSH2 region, demonstrating the GSH2 to be safe without affecting transcription of neighboring protein\-coding genes, and successful production and delivery of the KI transgene expression product into the host circulation, resulting in a partial neutralization of TTX\. Moreover, Good Manufacturing Practice \(GMP\) compliant methods for the preparation, production, storage, and administration of cryopreserved human hookworm are available[63](https://www.nature.com/articles/s41467-026-73447-9#ref-CR63)\. While genetic modification of the human microbiota has attracted considerable interest as a novel drug delivery strategy, the inability of most introduced bacterial species to persist long\-term in the gut remains a significant limitation\. In contrast, human hookworms readily establish following experimental infection and are exceptionally long\-lived, making them a stable and persistent pharmaceutical biofactory for the production and delivery of therapeutics of interest\.
Taken together, this transgenic human hookworm platform highlights a promising approach in biotechnology that has the potential to significantly advance how we conceptualize disease treatment and prevention\. Technologically, it also constitutes a notable advance in functional genomics for hookworms and helminths more broadly\.
## Methods
### Ethical statement
Studies using Syrian golden hamsters were carried out under protocols approved by Washington University School of Medicine \(WUSM; 20\-0323\) Institutional Animal Care and Use Committees \(IACUC\)\. All housing and care of laboratory animals conformed to the National Institutes of Health \(NIH\) Guide for the Care and Use of Laboratory Animals in Research \(see 18\-F22\) and all requirements and all regulations issued by the United States Department of Agriculture \(USDA\), including regulations implementing the Animal Welfare Act \(P\.L\. 89–544\) as amended \(see 18\-F23\)\. Roles and responsibilities were agreed upon amongst collaborators ahead of the research\. All unique materials are readily available from the authors or from standard commercial sources as detailed below\.
### Computational identification of Genomic Safe Harbors \(GSHs\)
A bioinformatic analysis utilizing available gene expression and genomic annotation data for*A\. ceylanicum*was carried out to identify optimal genome safe harbor \(GSH\) regions[11](https://www.nature.com/articles/s41467-026-73447-9#ref-CR11)\. The analysis was carried out to identify genomic regions that i\) contain putative promoter sequences associated with very high gene expression levels in bulk RNA\-seq data, increasing the likelihood that existing promoter and enhancer sequences in the region will facilitate the high expression of target genes, ii\) are upstream of genes with high gene expression across the life cycle, which reduces the probability of the genomic region being unavailable for transcription due to histone binding and iii\) have sufficiently long intergenic regions \( \> 2 kb\) to facilitate transgenesis without disrupting native gene expression, iv\) the sequence to be unique in the genome, v\) to contain a terminal[protospacer adjacent motif](https://www.sciencedirect.com/topics/biochemistry-genetics-and-molecular-biology/protospacer-adjacent-motif)\(PAM\) necessary for targeting by the sgRNA\-Cas9[ribonucleoprotein](https://www.sciencedirect.com/topics/biochemistry-genetics-and-molecular-biology/ribonucleoprotein)complex \(RNP\), and vi\) are situated \>2 kb from the nearest predicted coding region, following previously established guidelines[28](https://www.nature.com/articles/s41467-026-73447-9#ref-CR28)\. An available*A\. ceylanicum*genome assembly[64](https://www.nature.com/articles/s41467-026-73447-9#ref-CR64)\(PRJNA72583\) and annotation[65](https://www.nature.com/articles/s41467-026-73447-9#ref-CR65)were downloaded from WormBase Parasite[66](https://www.nature.com/articles/s41467-026-73447-9#ref-CR66)\. For the bioinformatic analysis, shorter gene IDs were assigned to replace the existing long gene IDs, to facilitate input into bioinformatic software \(original and assigned gene IDs are provided in Supplementary Data[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\)\. Specifically, the cumbersome gene IDs in the*A\. ceylanicum*annotation used \(e\.g\., “*ANCCEYDFT\_Contig1\-pred\_gff\_fgenesh\-gene\-5\.2\-mRNA\-1*”[65](https://www.nature.com/articles/s41467-026-73447-9#ref-CR65)\) have here been replaced with shorter IDs starting with “*ACEY\_*” and numbered with six digits after sorting the original gene IDs alphabetically \(e\.g\., “*ACEY\_000382*”\)\. In a newer*A\. ceylanicum*annotation[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17), produced since our GSH molecular analysis was performed for this study, “*ACEY\_*” gene IDs with five digits, a dash, and an isoform number are used \(e\.g\.,*ACEY\_11814\-1*\)\. To identify intergenic sequence motifs associated with highly expressed genes, up to 2000 bp of the 5’ sequence upstream of the 5’ untranslated region \(UTR\) or the first coding exon \(if no UTR was annotated\) was extracted for each gene\. Less than 2000 bp of upstream sequence was extracted for genes when either \(i\) the genomic sequence contig that the gene was on ended before 2000 bp, or \(ii\) another gene on either strand overlapped the upstream sequence, at which point the upstream sequence was terminated\. Using this approach, 2000bp sequences were retrieved for 79\.4% of genes, and at least 1000 bp were retrieved for 87\.1% of genes\. Transcriptomic RNA\-seq reads collected across the*A\. ceylanicum*life cycle were retrieved from a previous report[14](https://www.nature.com/articles/s41467-026-73447-9#ref-CR14), trimmed of adapters using Trimmomatic v0\.36[67](https://www.nature.com/articles/s41467-026-73447-9#ref-CR67), and mapped to the hookworm genome using the STAR aligner[68](https://www.nature.com/articles/s41467-026-73447-9#ref-CR68)\(v2\.7\.10a; 2\-pass mode, basic\)\. These included eight L3 stage larvae samples \(one 0\-hour, two 24\-hour, two 48\-hour and three 72\-hour distinct biological replicates\), three L4 larvae samples \(one 72\-hour, one 8\-day female, and one 8\-day distinct male biological replicate\), and eight adult samples \(four 16\-day male and four 16\-day female distinct biological replicates\)\. One adult male intestine RNA\-seq sample[34](https://www.nature.com/articles/s41467-026-73447-9#ref-CR34)was also mapped and quantified for the prioritization of signal peptide sequences\. Read fragment counts per gene per sample were quantified using featureCounts v1\.5\.1[69](https://www.nature.com/articles/s41467-026-73447-9#ref-CR69), and relative gene expression levels \(fragments per kilobase per million reads mapped; FPKM\) were calculated\. The average expression level for each gene across the*A\. ceylanicum*life cycle was calculated and ranked based on the average of the log10expression values\.*A\. ceylanicum*proteins were assigned functional annotations using results from InterProScan v5\.42[70](https://www.nature.com/articles/s41467-026-73447-9#ref-CR70)and GhostKOALA v2\.2[71](https://www.nature.com/articles/s41467-026-73447-9#ref-CR71)\. Potentially secreted proteins were identified using SignalP v5\.0[19](https://www.nature.com/articles/s41467-026-73447-9#ref-CR19)for signal peptides and transmembrane domains\. Also, reciprocal best\-hit BLAST matches between*A\. ceylanicum*,*A\. caninum*and*Necator americanus*were identified, to determine matches to genes confirmed by proteomics of ESPs in those hookworms[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17),[21](https://www.nature.com/articles/s41467-026-73447-9#ref-CR21)\.
### Motif enrichment and expression analysis to prioritize GSH sequences
The motif enrichment tool STREME[16](https://www.nature.com/articles/s41467-026-73447-9#ref-CR16)was used to identify enriched sequences among the 5’ upstream sequences \(maximum 2000 bp\) from the top 1% \(GSH1\) and the top 2% \(GSH2\) of expressed genes across the life cycle, compared to the bottom 50% lowest\-expressed genes\. The motif annotation tool TOMTOM[18](https://www.nature.com/articles/s41467-026-73447-9#ref-CR18)was used to match the top\-enriched motifs to putative transcription factors from the JASPAR database[25](https://www.nature.com/articles/s41467-026-73447-9#ref-CR25)\. FIMO[72](https://www.nature.com/articles/s41467-026-73447-9#ref-CR72)was used to identify all instances of the motifs in the 5’ upstream regions of all genes \(*P*≤ 10−5threshold for a match, according to FIMO[72](https://www.nature.com/articles/s41467-026-73447-9#ref-CR72)output\), and then STREME[16](https://www.nature.com/articles/s41467-026-73447-9#ref-CR16)was rerun on the genes containing the target motifs, to identify enriched “secondary” motifs significantly associated with the primary target motifs\. In the target GSH regions, these secondary motifs were used to define potential promoter boundaries to be avoided for transgene insertion\. The genes downstream of the target motifs were prioritized based on \(i\) high levels of gene expression across the life cycle, ii\) presence of a signal peptide for secretion[19](https://www.nature.com/articles/s41467-026-73447-9#ref-CR19), \(iii\) detection in*A\. ceylanicum*adult E/S products[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)and \(iv\) having more than 3000 bp of available 5’ upstream sequence, facilitating targeting the insertion of transgenes while minimizing the potential for disruption of promoter regions\. Supplementary Fig\.[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)summarizes the overall procedure for target gene identification and prioritization, while Supplementary Data[2](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)and[3](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)for GSH1 and GSH2, respectively, provide all motif sequences and positional nucleotide frequencies\.
For GSH1, the gene identified as*ACEY\_000382*was named*maker\-ANCCEYDFT\_Contig1\-pred\_gff\_fgenesh\-gene\-5\.2*in the version of the annotation used[65](https://www.nature.com/articles/s41467-026-73447-9#ref-CR65), and*ACEY\_11814\-1*in the most recent*A\. ceylanicum*annotation[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\. There were 9,078 bp of sequence preceding*ACEY\_000382*on the genome assembly and annotation used for the GSH prioritization analysis, but the most recent annotation[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)identified an additional highly expressed gene in the upstream intergenic region \(*ACEY\_11813\-1*\), shrinking the upstream GSH region to 3221 bp\.
For GSH2, the gene identified as*ACEY\_002225*was named*maker\-ANCCEYDFT\_Contig13\-pred\_gff\_snap\-gene\-4\.9*in the version of the annotation used[65](https://www.nature.com/articles/s41467-026-73447-9#ref-CR65), and*ACEY\_09100\-1*in the most recent*A\. ceylanicum*annotation[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\. There were 12,966 bp of sequence upstream of*ACEY\_002225*on the genome assembly/annotation used for the analysis, with the updated annotation[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)identifying a previously unannotated gene \(*ACEY\_09099\-1*\) 4,134 bp upstream of the gene\.
### Guide RNA and homology arm identification
The top two genome safe harbor regions \(GSH1 and GSH2\) were considered sites for CRISPR\-catalyzed KI of the gene of interest and its regulatory elements\. The sequences lacking repeats, low complexity, primary motif, secondary motif, and low\-confidence sequences associated with the primary motif of the GSH region were considered for generating gRNA\. Within these regions, we used the online tool CHOPCHOP[27](https://www.nature.com/articles/s41467-026-73447-9#ref-CR27)to identify sets of prioritized guide RNA \(gRNA\) sequences with protospacer adjustment motifs \(PAMs; “NGG” sequences\) among the upstream sequences for both*ACEY\_000382*\(GSH1: unidirectional two overlapping gRNAs: \(2 and 7, numbered based on the rank in CHOPCHOP\) and*ACEY\_002225*\(GSH2: unidirectional two overlapping gRNAs, 5 and 13; Supplementary Note[2](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)indicates the gRNA sequences and Supplementary Note[3](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)indicates genomic positions of the gRNAs\)\. The donor sequences also included 600 bp of homology arms complementary to the genomic DNA of either side across the programmed cleavage site of overlapping gRNAs\. The homologous sequences were considered after leaving 12 bp sequence from the cut site on either side\. All gRNA sequences were ordered from Integrated DNA Technologies \(IDT\)\.
### Hamster infection with infective\-stage A\. ceylanicum larvae
Syrian Golden Hamsters \(*Mesocricetus auratus*; Inotiv, HsdHan®:AURA strain\) are sourced from a colony established in 1994 with stock originating from Zentralinstitut fur Versuchstiere, producing an average litter size of 9\.0 and \(Hannover, Germany\)\. The “Han®” strain is a registered trademark of Inotiv\. Hamsters were healthy and weighed 50\-60 grams upon arrival\. Hamsters were housed in cages individually at ambient room temperature \(68\-72 °F\) and humidity \(range 30\-70%\), and fed LabDiet 5053 \(PicoLab® Rodent Diet 20, irradiated\)\. Only male hamsters were used for all experiments, since female hamsters are ~5\-fold less susceptible to*A\. ceylanicum*infection[73](https://www.nature.com/articles/s41467-026-73447-9#ref-CR73)\. Hamsters were randomly selected for experimentation, and for each experiment, the minimum number of hamsters required to produce the necessary samples \(eggs, worms, or serum\) was used\.
Male Syrian hamsters \(*Mesocricetus auratus*\), 21 to 28 days old, were infected with infective third\-stage larvae \(iL3\) of*A\. ceylanicum*by oral gavage[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)and in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health \(NIH\)\. Freshly isolated iL3s were counted and distributed in individual Eppendorf tubes, and 80 to 100 iL3s were resuspended per tube in 200 µl PBS[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\. A curved 18 G needle was used to gavage the hamsters\. Hamsters were manually restrained by grasping the loose skin at the scruff of the neck with the thumb and forefinger to immobilize the head and torso\. Using the needle held next to the animal, the distance from the tip of the nose to the last rib on the left side was measured\. Holding the animal with the nose pointing up, the gavage needle was inserted into the mouth over the tongue and directed toward the esophagus on the left side of the throat, gently pressing the needle on the back of the mouth and allowing the needle to slowly advance to the measured distance into the stomach\. Once the needle was properly positioned, the plunger was gently depressed to dispense ~200 µl of resuspended iL3s into the lumen of the stomach\. After delivery of the inoculum, the gavage needle was slowly withdrawn, and the hamster returned to its cage\. After the gavage, hamsters were observed for any difficulty breathing or for bleeding from the mouth or nose for at least 5 minutes before returning them to the vivarium\.
### Maintenance of the A\. ceylanicum life cycle in golden Syrian hamsters
At 14 days post\-infection, hamsters were placed on a wire at the bottom of cages lined with moist cardboard to facilitate fecal collection\. Mature eggs were extracted from these feces and processed for hatching to the iL3 stage using the Baermann funnel technique[74](https://www.nature.com/articles/s41467-026-73447-9#ref-CR74)for subsequent rounds of infection\. Alternate egg isolation methods for Neon protocol optimization and transfection experiments are detailed below in another section\. Hamsters were euthanized at 20 to 22 days after infection by CO2asphyxiation using compressed CO2gas as a source of carbon dioxide, allowing for the controlled inflow of gas to the induction chamber\. Hamsters to be euthanized were placed in a clean transparent chamber, and 100% CO2was introduced at a fill rate of 70% of the chamber volume per minute, to achieve a balanced gas mixture with the existing air in the chamber to fulfill the objective of rapid unconsciousness with minimal distress to the animals[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\. The expected time to unconsciousness was usually within 3 to 5 minutes\. Each hamster was monitored for lack of respiration and faded eye color\. CO2was maintained for a minimum of 1 minute after observing these signs to avoid unintended recovery upon exposure to natural CO2concentrations\. Upon completion of the procedure, death was confirmed by ascertaining cardiac and respiratory arrest and noting the animal’s fixed and dilated pupils\. Following euthanasia, the small intestine was removed and opened longitudinally and placed in incomplete RPMI 1640 medium \(Gibco\) in 100 mm petri dishes at room temperature \(RT\) for 1–2 hours\. During this incubation, adult worms actively migrated out of the intestinal tissue into the medium\. The intestinal tissue was then removed, and the remaining worms were gently detached from the mucosal lining using a fine nail brush\. The collected worms were washed three to five times with 1× PBS containing 5× Antibiotic\-antimycotic to avoid contamination\. Worms were then transferred to complete RPMI 1640 medium supplemented with 10% heat\-inactivated fetal bovine serum \(Gibco, Cat \#16140\-071\), 25 mM HEPES \(Corning, Cat \#25\-060\-CI\), 8\.9 mM sodium bicarbonate \(Corning, Cat \#25\-035\-CI\), and 1× Antibiotic\-Antimycotic\. Worms were maintained at 37 °C in a 5% CO₂ humidified incubator\. Worms were kept in sterile petri dishes and used for experiments on the same day or kept overnight for next\-day experiments\. For experimental treatments, worms were distributed into 24\-well tissue culture plates with 1 ml of supplemented RPMI medium per well\. Following electroporation \(when applicable\), worms were incubated in the same complete RPMI medium at 37 °C in a CO₂ incubator for 24 hours for recovery\. The following day, the medium was replaced with serum\-free RPMI containing 25 mM HEPES, 8\.9 mM NaHCO₃, and 1× Antibiotic\-antimycotic \(no FBS\), and worms were maintained in this serum\-free medium for up to 3–4 days\. Non\-responsive to touch \(likely dead\) worms were removed daily\. For evaluations, worms were washed thoroughly in 1× PBS and transferred into 1\.5 ml microcentrifuge tubes\. Depending on downstream applications, worms were either processed immediately for DNA or RNA isolation or flash\-frozen and stored at −80 °C\.
### Excretory\-Secretory Protein \(ESPs\) collection
ESPs were collected for WT and transgenic hookworms, following established protocls[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\. Adult worms were harvested from the small intestine of hamsters 18–22 dpi with wild\-type and transgenic iL3s, using the method described above\. The worms were washed three times in 1× PBS containing 5× antibiotic\-antimycotic\. They were cultured in complete RPMI media in triplicate at a density of 20 worms per well in 24\-well plates and incubated at 37 °C with 5% CO₂ for 4 hours\. After this pre\-incubation, the media were discarded to eliminate contamination from eggs, dead worms, and hamster tissue debris\. The worms were gently rinsed twice in PBS with antibiotics before transfer to 2 ml of serum\-free RPMI medium containing 2% antibiotic\-antimycotic and incubated again at 37 °C and 5% CO₂\. Over the next 3 days, 1\.5 ml of culture supernatant was carefully removed every 24 hours without disturbing the worms and replaced with fresh serum\-free medium\. The plates were returned to the incubator after each change\. Collected media were centrifuged at 4000 ×*g*for 30 min, 4 °C to minimize the contamination from eggs and dead worms, supernatant collected and stored at −80 °C\. After 72 hours, the worms did not exhibit visible damage or loss of motility, indicating that the ESPs collected reflected normal physiological activity of otherwise healthy*worms*\. At the end of the 72\-hour collection period, the stored ESP\-containing supernatant was slowly thawed on wet ice and concentrated using a centrifugal device with a 3 kDa molecular weight cutoff membrane \(Pall Corporation, Cat \#25\-035\-CI\)\. These concentrated ESPs were buffer exchanged by dialysis against PBS, protein content quantified using the BCA kit \(ThermoFisher, Cat \#23227\), and stored at −80 °C\.
### Chromosomal cleavage of the CRISPR targeted site \(GSHs\)
The efficacy of overlapping gRNAs \(IDT\), delivery method \(Bio\-Rad Electroporation vs Lipofection\), and comparison of two GSH regions was analyzed by comparing editing efficiencies with CRISPResso2[75](https://www.nature.com/articles/s41467-026-73447-9#ref-CR75)\. The hookworms were washed with serum\-containing medium \+ 2x Antibiotic\-Antimycotic after 3–4 hours of isolation and were again resuspended in the complete RPMI medium and kept at 37 °C with 5% CO2overnight\. The next day, 120 min before electroporation or lipofection, the worms were transferred into incomplete RPMI medium\. After 120 min, nutritional deprivation and worms more actively ingest the CRISPR reagents in the complete RPMI containing RNP complexes\.
The genome editing efficiency experiment was planned in three groups of adult worms \(*n*= 10 per group\) for each of the two GSH regions \(GSH1 and GSH2\)\. Group one \(control\) was not treated with the CRISPR RNP complex, while the second group was treated with the two unidirectional overlapping gRNAs\-Cas9 RNPs\. The RNP complexes were prepared by incubating each gRNA of the respective set with the Cas9 \(IDT, Cat \#10007807\) in equimolar concentration \(1 µM: 1 µM\) for 15 min in 20 µl Opti\-MEM medium \(Gibco, Cat \#31985\-062\), and then the overlapping gRNA\-Cas9 complex was pooled and mixed gently \(RNP complex\) without the formation of any bubbles, up to a total volume of 100 µl with Opti\-MEM Medium\. The RNP complex \(100 µl\) of overlapping gRNAs was mixed gently with the 10 adult*A\. ceylanicum*worms re\-suspended in 100 µl of Gene Pulser buffer \(Bio\-Rad, Cat \#1652676\)\. The adult*A\. ceylanicum*worms were incubated with RNP complex and Gene Pulser buffer for 30 minutes and then were electroporated using the Bio\-Rad electroporator under optimized*A\. ceylanicum*adult survival parameters \(Time Constant, 125 V, 20 milliseconds, and with two pulses\)\. Post electroporation, worms were kept at 37 °C in a CO2incubator for 72 hours\. Worms remained immobile for approximately 20–24 hours\. After electroporation, motility gradually resumed\. They were further maintained for an additional 48 hours\. to allow stabilization of gene incorporation\. Following incubation, adult worms were washed thrice with 2 ml of PBS before genomic DNA was isolated\. Genomic DNA was isolated using the DNeasy tissue and blood DNA isolation kit \(Qiagen, Cat \#69504\) and was stored at \-20 °C\. The genomic DNA used for the PCR amplification of the targeted region using a different set of primers depending upon the GSH \(Supplementary Note[4](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. After performing the experiment, the PCR amplicon of the target region of a specific size was purified using a PCR cleanup kit \(Invitrogen, Cat \#01318547\)\. The purified product was used as a template for PCR using the UDI index primer for Illumina sequencing\. The amplicon of the expected size was gel eluted and quantified using Nanodrop spectrometry and submitted for Illumina sequencing\. All primers were ordered from Integrated DNA Technologies \(IDT\)\.
### Lipofection
Micelles were prepared by diluting 9 µl RNAiMAX reagent \(Invitrogen, Cat \#13\-778\-100\) in 150 µl Opti\-MEM medium and incubating for 10 minutes at room temperature\. Then, 100 µl of the RNP complex made exactly the same way as the adult*A\. ceylanicum*electroporation protocol was combined with 150 µl of the diluted RNAiMAX micelles and was incubated for 20 min at room temperature \(RT\)\. Freshly isolated adult*A\. ceylanicum*worms were transferred to incomplete RPMI medium for 2\-3 hours before lipofection with RNP complex micelles\. After 2–3 hours, the worms were transferred into complete RMPI medium and treated with the RNP complex micelles and mixed gently\. The lipofected adult worms were maintained at 37 °C, 5% CO2for 72 hours\. At the end of the experiment, the adult worms were washed thrice with 2 ml of PBS, and the genomic DNA was isolated and sequenced as described above for electroporation\.
### Sequencing and analysis of chromosomal cleavage targeted site \(GSHs\)
Amplicon libraries were sequenced on Illumina’s NovaSeq 6000 S4, targeting 15 million read pairs per library\. Paired\-end sequencing reads \(2 × 150 bp\) were adapter/quality trimmed using trimmomatic v0\.39[67](https://www.nature.com/articles/s41467-026-73447-9#ref-CR67)and then merged using FLASH v1\.2\.11[76](https://www.nature.com/articles/s41467-026-73447-9#ref-CR76)\. The resulting amplicon sequences were analyzed for genome editing events using CRISPResso2 v2\.2\.12[75](https://www.nature.com/articles/s41467-026-73447-9#ref-CR75)\. A reference amplicon sequence corresponding to the targeted genomic locus in GSHs and the sgRNA sequences \(excluding the PAM sequence\) used in each experiment were provided to CRISPResso2 to infer the predicted position of editing activity\. The preprocessed amplicon reads were aligned to the reference sequence with the Needleman\-Wunsch algorithm, a global sequence alignment algorithm, and sequence modifications such as insertions, mutations, and deletions were quantified using the default quantification window size of 1 bp \(i\.e\., 1 bp on each side of the predicted cleavage position for a total length of 2 bp\)\. Finally, genome editing outcomes and efficiencies were estimated using CRISPRessoCompare by comparing the rate of sequence modifications in the experimental and the control groups\.
### Plasmid transformation and ESP collection for signal peptide testing
A pcDNA3\.1 plasmid containing an*A\. caninum*signal peptide \(Acan\-Asp2\) before a transgene was generously provided by Dr\. Mark Pearson of the Australian Institute of Tropical Health and Medicine[77](https://www.nature.com/articles/s41467-026-73447-9#ref-CR77),[78](https://www.nature.com/articles/s41467-026-73447-9#ref-CR78)\. This original plasmid DNA was cloned into pcDNA5/FRT/TO, a 5\.1 kb inducible expression vector designed to express and secrete recombinant proteins in mammalian cells, which included a CMV \(human cytomegalovirus\) promoter and an enhanced green fluorescent protein \(EGFP\) tag antibody engineered onto the C\-terminus of the transgene\. Signal peptide protein sequences for four other*A\. ceylanicum*target genes of interest \(as shown in Supplementary Fig\.[7](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\) were identified using SignalP v5\.0[19](https://www.nature.com/articles/s41467-026-73447-9#ref-CR19), and their corresponding coding sequences replaced the Acan\-Asp2 SP[32](https://www.nature.com/articles/s41467-026-73447-9#ref-CR32)in the expression vector using restriction sites strategically placed at the ends of the insert DNA fragments and within the restriction sites of the vectors\.*E\. coli*DH5α competent cells were transformed with the recombinant plasmids by heat shock\. Briefly, 50 µl of*E\. coli*DH5 competent cells were added to the ligation mixture, and the mixture was incubated on wet ice for 25 min, then placed in a 42 °C heat block for 30 sec\. Following ~5 min on ice, 200 µl of SOC recovery media \(Invitrogen, Cat \#15544034\) was added to the tubes, and the cells were allowed to recover at 37 °C with shaking at 900 rpm for 30 min\. Cells were plated on LB\-agar plates containing 100 µM carbenicillin and incubated at 37 °C overnight\. Plasmid DNAs were purified from 5 ml*E\. coli*DH5α overnight culture using the Invitrogen PureLink HiPure Plasmid Isolation Kit \(Invitrogen, Cat \#K210002\)\. Sanger DNA sequencing \(Genewiz, South Plainfield, NJ\) confirmed the identity of the inserted genes in the plasmids\.
The adherently growing human embryonic kidney cell line[79](https://www.nature.com/articles/s41467-026-73447-9#ref-CR79)\(HEK 293 T, Thermo Fisher Cat \#R71007; derived from human female embryonic kidney tissue\) was seeded in 6\-well plates at a density of ~2\.67 × 105cells per ml and cultured in a basic medium: Dulbecco’s modified Eagle’s medium \(DMEM; Gibco, Cat \#11965\-084\) supplemented with 10% \(v/v\) heat activated fetal bovine serum \(HI\-FBS, Gibco, Cat \#16140\-071\), 2 mM glutamine, 1x penicillin\-streptomycin and 1x non\-essential amino acids at 37 °C with 5% CO2\. After 24 hours, the cells showed 85% confluency and were transfected with 2 µg of plasmids using X\-tremeGENE HP DNA transfection reagent \(Millipore\-Sigma, Cat \#XTGHP\-ROG\) according to the manufacturer’s instructions\. Plasmids were diluted with 200 µl of opti\-MEM \(ThermoFisher Scientific, USA, \#31985070\), and a 3:1 ratio of reagent to DNA was used to determine the amount of X\-tremeGENE reagent to add\. HEK 293 T cells were not authenticated or tested for mycoplasma contamination\. After 4 hours of incubation of HEK 293 T cells with the transfection reagent and DNA complex, 1 µl of 1 mg/ml doxycycline was added\. The supernatant was collected every 16 hours, and fresh media was added at each collection\. Cells and culture media from 72 hours post\-transfection cells were collected for subsequent experiments\. All produced DNA constructs are available for reuse upon request to the corresponding author\.
Isolation of adult hookworms from hamsters was performed following established protocols[20](https://www.nature.com/articles/s41467-026-73447-9#ref-CR20)\. Freshly recovered synchronized adult*A\. ceylanicum*were cultured in FBS\-rich media: RPMI\-1640 media \(Gibco, Cat \#21870\-076\) supplemented with 25 mM HEPES \(Corning, Cat \#25\-060\-C1\), 25 mM NaHCO3\(Corning, Cat \#25\-035\-C1\), 1x penicillin\-streptomycin \(ThermoFisher Scientific, Cat \#15140122\), and 10% \(v/v\) FBS \(Gibco \# A4766801\) at 37 °C with 5% CO2\. After 20 hours, the worms were gently washed in 1x PBS \(Corning, Cat \#21\-040\-CMR\) containing 5x Antibiotics/Antimycotics \(Sigma Aldrich, Cat \#A5955\) and starved for 4 hours \(to facilitate the ingestion of the plasmids into their gastrointestinal system before electroporation\) in FBS\-free media: RPMI 1640 Media \(Gibco, Cat \#21870\-076\), 1x Glutamax \(Gibco, Cat \#35050\), 1x Streptomycin\-Penicillin \(ThermoFisher Scientific, Cat \#15140122\) at 37 °C with 5% CO2\. Afterward, 30 worms in each condition were incubated in an electroporation complex containing 80 µl Gene Pulser Electroporation Buffer \(Bio\-Rad, Cat \#1652676\)\. Worms were electroporated with or without 10 µg of plasmid construct at 125 V for 20 ms \(x2\) using Gene Pulser Xcell Electroporation Systems \(Bio\-Rad, Cat \#1652660\)\. Following electroporation, the worms and electroporation complex were transferred to 24\-well plates, and 2 ml FBS\-rich media was added to each condition\. Every 24 hours, over 3 days, at least 1\.8 ml of culture supernatant was aspirated from each well with minimal disturbance to the hookworm\. Following established protocol[20](https://www.nature.com/articles/s41467-026-73447-9#ref-CR20), each well was replaced with fresh serum\-rich media and transferred to the incubator\. At the end of 72 hours of culture medium collection, the worms were gently washed in 1x PBS with 5x Antibiotic/Antimycotic, and the culture supernatant containing ESPs was gradually thawed in ice\. Approximately 5\.4 ml of culture supernatant was recovered from 72 hours of media collection per condition\. The crude ESP supernatant was centrifuged at 7000 ×*g*at 4 °C for 30 min before concentration and transferring to a new tube\. Using this technique prevented laid eggs and remaining tissue from hamsters or dead worms from clogging the centrifugal filter\. The ESP supernatant was concentrated using 20 ml Pall Macrosep Advance Centrifugal Devices with a defined 3 kDa cutoff membrane \(Pall Life Sciences, Cat \#MAP003C37\)\. Briefly, 13 ml of culture supernatant was placed into the concentrator sample chamber and capped\. The concentrator was then placed into the bucket rotor that contained a 20 ml tube\. Concentration by 3 kDa Pall membrane was performed at 5000 ×*g*at 4 °C until the retention volume was reduced to 0\.5 ml\.
### Western blot analysis for signal peptide testing
Transfected HEK 293 T cells were washed with 1x PBS to remove the residual FBS\-rich medium\. The cells were harvested in 1x PBS and centrifuged to collect the pellet 500 ×*g*for 5 min\. 200 µl of 2x sample buffer \(made from Bio\-Rad XT Sample Buffer 4X, Bio\-Rad, Cat \#1610791\) with 5% \(v/v\) 2\-Mercaptoethanol \(BME, Sigma\-Aldrich, Cat \#60\-24\-2\) was added to the cell lysate in each signal peptide condition\. The mixtures/cells were lysed using a Q700 QSONICA Sonicator \(M2 Sci, Cat \#Q700\) set at an amplitude of 60 with 30 sec on pulse time and 30 sec off pulse time\. Lysed cells were heated at 95 °C for 5 min, and the supernatant was centrifuged to collect any cells at the bottom of the collection tube\. Supernatant was transferred to a new tube to add 200 µl of 3x sample buffer with 5% BME to 100 µl of each media supernatant\. The mixture was sonicated and re\-heated to 95 °C for 5 min\. The HEK 293 T cell supernatant and worm lysate ESPs \(collected as described above\) were quantified using a Pierce Coomassie Plus Bradford Protein Assay \(ThermoFisher Scientific, USA, Cat \#A55866\)\. Separately, 5 µl of each sample was then added to 395 µl of 1× Bradford dye in a 96\-well plate\. Following 5 min of incubation at RT in the dark, the optical density \(OD\) at 595 nm was assessed by spectrophotometry \(Eon Microplate Spectrophotometer, Biotek Instruments, Agilent\)\. The protein concentrations of both ESP supernatant and lysate were estimated based on linear extrapolation from a standard curve established using dilutions 1 mg/ml bovine serum albumin \(BSA\) supplemented with 2% Antibiotics/Antimycotics \(Gibco; ThermoFisher Scientific, USA, Cat \#15240112\)\.
Approximately 50 μg of each sample was mixed with 2x concentrated Laemmli\-SDS buffer \(Bio\-Rad, Cat \#1610737\) with 5% 2\-mercaptoethanol \(Sigma\-Aldrich, Cat \#M6250\)\. The samples were heated at 95 °C for 5 minutes, then 50 µl of each sample was separately loaded into the wells of 12% Criterion XT precast gel \(Bio\-Rad, Cat \#3450123\)\. The precision protein standard \(Bio\-Rad, Cat \#1610374\) was loaded into the first well at 10 µl without dilution with an SDS sample buffer\. Electrophoresis was run for 60 min at 14 watts using 1x running buffer \(XT MES and Milli\-Q water, from the kit\)\. The filter papers were soaked in semi\-dry transfer buffer \(25 mM Tris, 192 mM glycine, and 10% \(v/v\) Isopropanol\), and the membrane was immersed in methanol for 20 min before transfer\. The proteins were transferred to the membrane by semi\-dry protein transfer at 25 V for 40 min\. The membrane was blocked in 5% milk for 60 min at RT, followed by incubation with an anti\-GFP \(Clontech, Cat \#632592\) probe at 4 °C overnight with shaking\. The membrane was then washed with 1x PBST buffer \(1X PBS and Tween 20, ThermoFisher Scientific, Cat \#28320\) for 10 min, and 5 min in PBS only\. The GFP sample was further incubated with the anti\-mouse \(GE Healthcare Life Sciences, Cat \#NA931\-1ML\) at 4 °C for 60 min and washed again\. The membrane was incubated in western blot substrate at a 1:1 ratio \(Stable Peroxide Substrate and Luminol/Enhancer Buffers\) and immediately visualized with Bio\-Rad Gel Doc EZ Imaging System \(Bio\-Rad, Cat \#1708270\)\. Blots were imaged in Azure Imaging Systems \(C600, Azure Biosystems\) under LED visible fluorescence detection mode\.
### Production of recombinant s16\-HuScFv
The*s16\-HuScFv*sequence was obtained from Chulanetra et al[13](https://www.nature.com/articles/s41467-026-73447-9#ref-CR13)for expression in ExpiCHO cells \(Thermo Fisher, Cat \#A29127; derived from female Chinese hamster ovary\)\. The open reading frame \(ORF\) consisted of the*ASP\-1*signal peptide, the*s16\-HuScFv*sequence, and a C\-terminal 6\-His tag\. cDNAs were synthesized with mammalian codon bias by Genscript and cloned into the pcDNA3\.1 plasmid by restriction cloning\. Plasmid was purified and introduced into ExpiCHO cells by lipofection using an ExpiFectamine CHO transfection kit \(Thermo Fisher, Cat \#A29130\) as per the manufacturer’s instructions \(performed by GenScript\)\. Recombinant protein was purified on an AKTA FPLC by immobilized metal affinity chromatography using His\-trap excel nickel column and buffer exchanged into tissue culture grade DPBS using Amicon ultra\-3 centrifugal concentrators and quantified using a bicinchoninic acid kit \(Thermo Fisher, Cat \#23225\)\. Recombinant protein was assessed for endotoxin using a Limulus Amoebocyte Assay \(Thermo Fisher, Cat \#Q32892\) and only used if endotoxin levels were less than 0\.5 endotoxin units per mg protein\. Wherever possible, endotoxin\-free plasticware was used\.
### Titration and Western blot
The recombinant s16\-HuScFv produced from CHO cells was used as a positive control\. To determine the minimum detectable protein concentration using a western blot with anti\-6x\-His, HRP antibody \(Invitrogen, Cat \#MA1\-21315, Lot \#3216088\), we aimed to quantify the 16\-HuScFv secreted by the transgenic hookworms\. Antibody validation is supported by the manufacturer’s ‘Advanced Verification’ data \(accessible via the Thermo Fisher product catalog page,[https://www\.thermofisher\.com/antibody/product/6x\-His\-Tag\-Antibody\-clone\-HIS\-H8\-Monoclonal/MA1\-21315](https://www.thermofisher.com/antibody/product/6x-His-Tag-Antibody-clone-HIS-H8-Monoclonal/MA1-21315)\), which includes Western blot testing confirming expected target molecular weights and a lack of cross\-reactivity with off\-target epitope tags \(e\.g\., HA\-tags\)\. The initial protein concentration was 600 µg/ml, which was diluted to 150, 75, 37, 18, 9, 4\.5, and 2\.25 µg/ml and 20 µl added in each well of the Bio\-Rad precast gel of 4–15% were run for 90 min at 120 V before setting the transfer of protein on PVDF membrane for another 60 min at 100 V in the Bio\-Rad wet transfer unit at 4 °C\. After the transfer, the membrane was blocked in 5% skimmed milk in TBST \(Tween 20 detergent in Tris\-buffered saline\) for 30 minutes\. The blot was incubated with anti\-6x\-His, HRP \(1:3000 dilution\) overnight at 4 °C\. The blot was washed three times for 10 min\| each with TBST before ECL treatment and chemiluminescent scanning in \(Gel doc, Azure Biosystem\)\.
### Preparation of donor construct with short \(50 bp\) homology arms
The 50 bp donor, used only for initial Neon electroporation protocol optimization, was prepared by PCR amplification from the*s16\-HuScFv*expression component of the expression plasmid pcDNA5\-TTX*s16\-HuScFv*\. Forward and reverse primers were designed with 50 bp of homology across the target*A\. ceylanicum*genomic sequence 12 bp away from the cut site of the gRNA\. These primers also included 15–20 bp sequence from pcDNA5\-TTX*s16\-HuScFv*, complementary to the cloning start site for the donor preparation\. The total sequence for these primers varies from 65\-70 bp \(50 bp from the homologous region and 15–20 bp of the plasmid priming sequence\)\. In an Eppendorf tube, 10 µM of primer \(either forward or reverse\) was incubated with 1 mM Bis\-PEG10\-NHS ester in 1× Borate buffer overnight at RT\. The reaction was stopped in the morning using 0\.2 M EDTA \(pH 8\)\. These primers were desalted using a Bio\-spin 30 column \(Bio\-Rad\)\. These desalted primers \(Forward/Reverse\) were used for the donor amplification from pcDNA5\-TTX*s16\-HuScFv*\. Once the donor was amplified from the pcDNA5\-TTX*s16\-HuScFv*by the above\-designed primer, these PCR amplicons were run through the gel, and the expected band was gel eluted\. The concentration of amplicon was measured, and 5 µg donor DNA was used for the electroporation\. In downstream experiments described below, longer 600 bp HAs were used\.
### Preparation of donor construct with long \(600 bp\) homology arms
The donor construct, CMV\-Asp1\-s16\-HuScFv 6xHis \-BGH with right and left homology arms of 600 bp length, was chemically synthesized and engineered by subsequently cloning into pcDNA3\.1\(\+\) vector by GenScript at the*Not*I enzyme \(NEB, Cat \#R3189S\) restriction site\. N\-terminal optimization[36](https://www.nature.com/articles/s41467-026-73447-9#ref-CR36)included modifying nucleotides 7 to 15 to introduce A/U\-biased hexanucleotide sequences and optimizing the corresponding amino acids at codons 3 through 5 \(e\.g\., utilizing the KN\-YI motif\) to prevent ribosomal pausing and abortive peptidyl\-tRNA drop\-off during early translation elongation\. Homology arms of 600 bp in length were used for all experiments besides the Neon electroporation protocol optimization\. DH5α strain*Escherichia coli*cells were transformed with the plasmid, positive colonies were selected on the LB ampicillin \(Gold Bio, Cat \#A\-301\-25\) plates, and the plasmid was isolated using the QIAGEN Plasmid*Plus*Maxi Kit \(Qiagen, Cat \#12163\)\. The purified plasmid was digested with*Not*I overnight at 37 °C, cleaved fragments separated by agarose gel electrophoresis, and the transgene\-bearing fragment \(3\.2 kb\) eluted using Zymoclean Gel DNA recovery kit \(ZymoResearch, Cat \#11\-301\) into 50 µl nuclease free water\. The concentration and purity of this double\-stranded DNA donor were established by spectrometry \(Nanodrop\) before storage at −20 °C\. All produced DNA constructs are available for reuse upon request to the corresponding author\.
### Evaluation of Neon transfection protocols using A\. ceylanicum eggs
Male Golden Syrian hamsters \(*Mesocricetus auratus*, 3–4 weeks old\) were infected orally with 80\-100 iL3s of*A\. ceylanicum*in 200 µl of PBS using an oral gavage needle\. After 18–20 days of infection, hamsters were euthanized, the small intestine was removed and opened longitudinally as described above\. Approximately 200–250 adult worms were collected and washed as above, and then cultured overnight in RPMI medium supplemented with 10% FBS \(Gibco\) and 1x antibiotic and antimycotic \(Gibco\) at 37 °C in 5% CO2in air to facilitate egg laying\. Eggs were collected by centrifugation at 2000 ×*g*, 2 min, room temperature \(RT\), supernatant was removed, and the eggs assayed to evaluate CRISPR transgene delivery performance of the Neon Electroporation system \(Invitrogen, Cat \#MPK5000\)\.
### Isolation of in vivo laid eggs from hookworm\-infected hamster feces
After 16 days p\.i\. with 80–100*A\. ceylanicum*iL3, hamsters were transferred to a cage with wire flooring for 18–24 hours to enable collection of feces\. Hookworm eggs were isolated from these feces according to established methods[80](https://www.nature.com/articles/s41467-026-73447-9#ref-CR80), with minor modification\. Briefly, feces were soaked in 13% NaCl solution \(Sigma\-Aldrich, Cat \#S9888\) \(two ml per gram of feces\) for 30 minutes at RT, after which the fecal slurry was homogenized with a spatula and then pelleted by centrifugation at 2000 ×*g*, for 5 minutes at RT\. Following centrifugation, the supernatant containing eggs was decanted into a beaker with an equal volume of distilled water\. This egg suspension was centrifuged at 2000 ×*g*, 5 min, the supernatant discarded, and the pellet containing eggs was resuspended in 10 ml sucrose \(17% in water\) \(Sigma\-Aldrich, Cat \#S7903\)\. Next, resultant suspension was clarified \(2000 ×*g*, RT\) and the supernatant was split into two tubes and filled with 15 ml of distilled water and centrifuged as above\. The pellet was resuspended in 4\.5 ml of distilled water containing 0\.5 ml of 6% hypochlorite \(Thermo Fisher, Cat \#SS290\-1\), gently rocked for 60 sec, and centrifuged at 800 ×*g*, 2 min, RT, and the pellet was washed three times with sterile double\-distilled water\. Finally, eggs were resuspended in 1 ml of PBS and placed on a 30 μm pore size sieve \(PluriSelect, Cat \#43\-50030\-03\) to wash and remove debris, and the eggs were collected above the mesh and counted\.
### Evaluation of Neon transfection protocols using A\. ceylanicum eggs
In vitro laid eggs were washed with Opti\-MEM media, then resuspended in 1\.0 ml Opti\-MEM media\. Eggs were counted and dispensed in equal numbers of eggs into each of 25 tubes\. To prepare the RNA\-guided Cas9 ribonucleoprotein \(RNP\) complex, the overlapping gRNA set \(2 overlapping gRNAs of GSH2; 5 and 13\) was diluted to a concentration of 1 µM in Opti\-MEM medium\. Simultaneously, Cas9 \(Alt\-R™ S\.p\. Cas9 V3, glycerol\-free \(IDT, Cat \#10007807\) protein was diluted to a final concentration of 1 µM in Opti\-MEM medium\. The diluted gRNA and Cas9 were mixed in an equal ratio \(50 µl:50 µl\) and incubated for 15 min at RT to form RNP complexes\. Following incubation, 30 µg of double stranded linear donor DNA \(50 bp HAs\) was added to the RNP complex\. For Neon electroporation, an equal volume of T\-Buffer was added to the eggs/RNP complex in each tube, which were then subjected to electroporation using 24 different protocols\. These varied in pulse voltage, pulse width, and pulse number on the Neon transfection system \(see Supplementary Fig\.[9](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. After electroporation, all 24 different Neon protocols were evaluated by PCR using different sets of primers \(Supplementary Note[5](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. In downstream experiments described below, longer 600 bp HAs were used\.
### Transfection of eggs with the transgene
Two hundred and fifty eggs each were used for the Mock \(control\) group \(no Cas9/RNP complex, no s16\-HuScFv Donor DNA\), the Donor only \(no Cas9/RNP complex, only with s16\-HuScFv Donor DNA\), and the experimental group samples \(with Cas9/RNP complex and s16\-HuScFv donor DNA\)\. Three biological replicates were generated for each group\. RNA\-guided Cas9 \(gRNA\.Cas9\) ribonucleoprotein \(RNP\) complex was prepared as described above\. After incubation, 6 µg of double stranded linear donor \(Asp1\-s16\-HuScFv\) was added to the gRNA/Cas9 RNPs\. The total volume was adjusted to 350 µl and was added to the 50 µl of eggs\. For Neon electroporation, an equal volume of T\-Buffer \(Neon Transfection system kit, Thermo Fisher, Cat \#MPK10096\) was added to the egg’s gRNA/Cas9 RNP resultant suspension and were electroporated using protocol 13 \(Pulse voltage: 1100 V, Pulse width: 20 milliseconds, Pulse number: 2\) of the Neon transfection system, and electroporated eggs used for genomic DNA isolation and evaluated for the integration of s16\-HuScFv transgene by PCR and ONT sequencing\.
### Collection of transgenic F0 adults and F1 eggs
Hookworm eggs isolated from hamsters and electroporated were transferred to 24\-well plates containing 1× PBS supplemented with 1% penicillin \(10,000 U per ml\) and streptomycin \(10 mg per ml; Sigma\-Aldrich\) and 5% amphotericin B \(250 μg per ml; Sigma\-Aldrich\) and placed in a CO2incubator at 28 °C to induce hatching to the L1 larval stage\. The following day, the L1 larvae were transferred to the nematode growth media \(NGM\) plates seeded with OP50 \(Lot \#053114\) to develop into the infective third\-stage larva \(iL3\) on NGM plates incubated at 28 °C at 5% CO2for 3 days\. Following incubation, iL3s were collected, washed with PBS, and maintained in BU buffer until used to infect hamsters\. The iL3s were counted, and each hamster was infected with 80\-120 iL3s, as above\. To evaluate the germline transgenesis, feces were collected from these infected hamsters at 18, 19, and 20 dpi\. On each collection day, F1 eggs were isolated as above and used for genomic DNA \(g\.DNA\) isolation, PCR, and ONT sequencing\. F0 adult worms were collected as described above from hamsters and g\.DNA was isolated from every single worm and evaluated for the integration of s16\-HuScFv transgene by PCR and ONT sequencing\. On\-target, programmed transgene KI was confirmed by amplification of the targeted genomic region, followed by amplicon sequencing analysis\.
### Genomic DNA isolation from electroporated eggs and adults
g\.DNA isolation from eggs was performed following the protocol for the DNeasy Blood & Tissue Kit \(QIAGEN\)\. Briefly, 180 μL ATL buffer and 20 μL proteinase K were added to the electroporated eggs, mixed by vortex, and incubated at 56 °C for 3 hours\. Subsequently, 200 μL AL buffer was added and incubated at 56 °C for 10 minutes\. Following incubation, 200 μL of ethanol \(96\-100%\) was added to the suspension and mixed\. The suspension was pipetted into a DNeasy Mini spin column, centrifuged at ≥ 6000 ×*g*for 1 min\. Subsequently, the spin column was washed with 500 μL of Aqueous Wash 1 \(AW1\) buffer \(Aqueous Wash\) and 500 μL of AW2 buffer\. DNA was eluted in 50 μL of nuclease\-free water and stored at −20 °C until use for PCR and ONT sequencing\. For g\.DNA isolation from a single adult*A\. ceylanicum*worm, the worm was ground in 25 μL of the Grinding Buffer \(80% Homogenization Buffer comprising 100 mM NaCl, 200 mM sucrose, 10 mM EDTA \(Corning, Cat \#46\-034\-Cl\), and 30 mM Tris pH 8\.0 \(Corning, Cat \#46\-034\-CM\) and 20% Lysis Buffer consisting of 250 mM EDTA pH 8\.0, 2\.5% SDS \(Invitrogen, Cat \#15553\-035\), and 500 mM Tris pH 9\.2\)\. The worm lysate was incubated at 65 °C for 30 minutes\. Following incubation, 7 µL of 8 M ammonium acetate \(Sigma\-Aldrich, Cat \#A1542\) was added to the lysate and incubated in wet ice for 60 minutes\. Subsequently, the lysate was clarified by centrifugation, 2500 × g for 10 min\. The supernatant was collected, and 100 μL of 100% ethanol was added and incubated at room temperature for 5 min\. Then, the mixture was centrifuged at 18,500 ×*g*, 15 min, 4 °C, and the supernatant removed\. The barely visible pellet was washed with 100% ethanol, air dried, and dissolved in 50 μL of 10 mM Tris and 1 mM EDTA \(TE\) and stored at \-20 °C until use for PCR and ONT sequencing\.
### Detection of transgenesis in F0 eggs, F0 adults, and F1 eggs by PCR
Detection of integration of the transgene into the genome of electroporated eggs \(F0 eggs\), F0 adults worms and F1 eggs at GSH2 was undertaken using PCR, using Phusion®High\-Fidelity DNA Polymerase \(NEB, Cat \#M0531S\) and informative primers; one primer set targeted the GSH2 using specific primers upstream \(5’up\_F3\) of the homology arms paired with primers \(CMV\_R\) specific for the*CMV*promoter region of donor DNA and primer \(s16 R\) specific for the s16\-HuScFv \(sequences in Supplementary Note[6](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. The PCR cycling conditions included the initial denaturation step at 98 °C for 5 minutes, followed by 30 cycles of denaturation at 98 °C for 30 seconds, annealing at 65 °C or 67 °C for 30 seconds, extension at 72 °C for 2 minutes, and final extension at 72 °C for 10 minutes\. The amplification products were sized by electrophoresis through agarose gel and visualized and documented under UV light \(Gel doc, Azure Biosystem\)\. The amplicon sizes expected at the 5’ integration site were 1,061 bp and 2365 bp using primer pairs 5’up\_F3 &CMV R and 5’up F3 & s16 R, respectively\. The integrity of g\.DNA from electroporated eggs \(F0 eggs\), F0 adult worms, and F1 eggs were confirmed by amplification of the*Rab11a/b*constitutive gene using Rab11a/b*F*& Rab11a/b R primers \(Supplementary Note[6](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. The amplified products from F0 eggs, F0 adult worms, and F1 eggs were eluted from the gel \(Zymoclean Gel DNA recovery kit\) and nucleotide sequences of amplicons determined using ONT sequencing \(Plasmidosaurus\)\. All primers were ordered from Integrated DNA Technologies \(IDT\)\.
### qRT\-PCR quantification of gene expression
Total RNA was isolated using Trizol reagent \(Invitrogen, Waltham, MA, USA\) from four independent experiments\. Each experiment has one biological replicate of approx\. ~20,000 eggs electroporated in the absence of the transgene construct \(mock/WT\), and one replicate with the s16\-HuScFv construct \(above; “transgenic”\. RNA was also isolated from two pools of WT and transgenic adults \(4 worms per replicate\), after first homogenizing using 1\.5 mm zirconium beads \(Benchmark scientific Cat \#D1032\-15\) using BeadBug6 \(Benchmark scientific, Cat \#D1036\)\. 200 µl of chloroform was added, followed by vigorous vortexing and incubation for 5 minutes\. After phase separation, the aqueous layer containing RNA was transferred to new tube, and RNA was precipitated using 500 µl of isopropanol and incubation at room temperature for 20 minutes\. Samples were centrifuged at 12,000 ×*g*for 10 minutes at 4 °C\. The supernatant was discarded, and the RNA pellet was washed with 1 ml of 75% ethanol, briefly vortexed, and centrifuged at 7500 ×*g*for 5 minutes at 4 °C\. Pellets were air dried for 5–10 minutes and resuspended in 35 µl of RNase\- free water, 15 µl stored at −80 °C for RNA sequencing, and the remainder employed as the template for cDNA synthesis and qRT\-PCR\. Transgenesis of eggs was confirmed by PCR as described above\. Three samples each from the WT and putatively transgenic eggs were selected for qRT\-PCR quantification of gene expression of genes of interest\. Residual genomic DNA was removed by DNase I digestion with the DNA\-free DNA removal kit \(Invitrogen, Cat no\-AM1906\)\. RNA was quantified by Qubit HS RNA assay \(Life Technologies\) and its integrity determined by RNA assay on 4200 Tapestation \(Agilent\)\. cDNA was synthesized using the readyScript cDNA synthesis kit \(Sigma, Cat no \- RDRT\) according to the manufacturer’s instructions\. qRT\-PCR reactions were performed using PowerUp™ SYBR™ Green Master Mix \(Applied Biosystems, Cat no\-A25742\) on QuantStudio™ 3 Real\-Time PCR System \(Invitrogen\)\.*Rab11a/b*served as representative endogenous reference genes\. The relative expression was calculated by using 2\-∆CTmethod, using*Rab11a/b*as the reference\. When no signal was detected, a CT value of 40 was used for calculations, representing the background detection limit\. Supplementary Note[6](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)lists the primers and their sequences employed for endogenous controls \(*BenA*and*Rab11a/*b\),*Acey\_002221*\(GSH upstream gene*\), Acey\_002225*\(GSH downstream gene\) and*s16\-HuScFv*\. Significant differences between WT and transgenic samples were assessed in two\-tailed*t*\-tests \(equal variance\), using the ∆CT values, before transformation to 2\-∆CTgene expression values\.
### RNA seq analysis
The three WT and transgenic egg RNA biological replicates with the best QC \(RIN \> 8\.6\) from the qRT\-PCR analysis described above underwent bulk RNA\-seq profiling\. cDNA libraries were prepared using PolyA selection, and processed cDNA was sequenced on the Illumina NovaSeq S4 platform \(paired\-end 150 bp reads\), generating an average of 17\.4 million read pairs per sample\. The raw RNA\-seq read files \(fastq\) are accessible on the NCBI Sequence Read Archive \(SRA[81](https://www.nature.com/articles/s41467-026-73447-9#ref-CR81), BioProject PRJNA1328670\), and read count statistics and SRA accession information per sample are provided in Supplementary Data[4](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\. Sequenced reads were mapped to the*A\. ceylanicum*genome assembly[15](https://www.nature.com/articles/s41467-026-73447-9#ref-CR15),[64](https://www.nature.com/articles/s41467-026-73447-9#ref-CR64)\(PRJNA72583\) and annotation[65](https://www.nature.com/articles/s41467-026-73447-9#ref-CR65), using tools and settings as described above for samples across the life cycle, in the “Database production for GSH analysis” section of the Methods\. DESeq2 \(v 1\.44\.0\)[42](https://www.nature.com/articles/s41467-026-73447-9#ref-CR42)was used to perform differential expression analysis between WT and transgenic samples \(*n*= 3 each, with batch effects for each WT\+transgenic pair\), with thresholds of FDR\-adjusted*P*≤ 0\.05 and \|Log2Fold Change \| ≥ 1 for significant differential expression\. The DESeq2 R script was retrieved from protocols\.io \([https://doi\.org/10\.21203/rs\.3\.pex\-2532/v1](https://doi.org/10.21203/rs.3.pex-2532/v1)\)\. Supplementary Data[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)provides read counts, normalized gene expression levels, and all DESeq2 differential expression statistics for all genes\.
### ELISA\-based assays for s16\-HuScFv quantification
ELISA was performed to quantify s16\-HuScFv levels in hamster serum\. Nunc ELISA plates were coated with 2 µg/ml anti\-His antibody \(ThermoFisher, Cat \#MA1\-21315, Lot \#3216088\) diluted in carbonate buffer \(15 mM NaHCO3, 30 mM NaH2CO2, and 0\.02% sodium azide, pH 9\.2\) and incubated overnight at 4 °C\. The next day, plates were washed three times with wash buffer \(1× Dulbecco’s phosphate\-buffered saline \(DPBS\), pH 7\.2, containing 0\.05% Tween\-20\) and then blocked with 300 µl of blocking buffer \(2% BSA in wash buffer\) for 2 hours at room temperature \(RT\)\. After blocking, plates were washed again and incubated with ESP collected from TG hookworm, undiluted hamster serum, or 1:10 diluted hamster serum for 2 hours at 37 °C\. For generating the standard curve, serial dilutions of recombinant purified s16\-HuScFv were prepared, and 100 µl of each dilution was added to the corresponding wells and incubated under the same conditions\. Plates were then washed with 200 µl/well of wash buffer three times for 5 minutes each on a shaker\. Subsequently, a 1:5000 dilution of HRP\-conjugated goat anti\-human IgG F\(ab’\)₂ fragment\-specific antibody \(Abcam, Cat \#109\-035\-097, Lot \#174499\) was added, and the plates were incubated for 60 min at RT\. Antibody validation is supported by the manufacturer’s “purity” and “antibody specificity” analysis in the product specifications for this lot \([https://www\.jacksonimmuno\.com/lots/000000174499](https://www.jacksonimmuno.com/lots/000000174499)\), which includes immunoaffinity chromatography purification, immunoelectrophoresis, and ELISA with positive and negative controls\. After incubation, plates were washed six times \(200 µl/well\) for 5 minutes each on a shaker\. Signal development was performed using 1\-Step ultra TMB\- ELISA substrate \(ThermoFisher, Cat \#34028\), and the reaction was stopped with 2 M H₂SO₄\. Optical density was read at 450 nm using a BioTek Eon Microplate Reader \(Midland, ON, Canada\)\.
### Tetrodotoxin neutralization assays
Murine neuroblastoma Neuro\-2a cells \(Merck, Cat ECACC 89121404\) were cultured in ATCC 30\-2003 Eagle’s Minimum Essential Medium \(EMEM\) containing Earle’s balanced salt solution, non\-essential amino acids, 2 mM L\-glutamine, 1 mM sodium pyruvate, and 1500 mg/L sodium bicarbonate\. The EMEM was supplemented with 10% heat\-inactivated fetal bovine serum \(Bovogen Biological, East Keilor, VIC\) and antibiotics \(10,000 U/ml penicillin and 10,000 µg/ml streptomycin; Gibco, 15140122; Thermo Fisher Scientific, Scoresby, VIC\)\. Cells were maintained at 37 °C, 5% CO2\. For passaging, spent medium was removed, and adherent cells were rinsed with 10 ml of pre\-warmed PBS \(calcium\- and magnesium\-free\)\. Detachment was induced by incubating cells with ~3 ml of pre\-warmed TrypLE™ Express Enzyme \(1X, no phenol red; Gibco\) for 5 to 15 minutes at 37 °C\. Cells were resuspended in 7 ml of culture medium, collected into a 15 ml tube, and centrifuged at 130 ×*g*for 7 minutes at room temperature\. The supernatant was discarded, and the cell pellet was resuspended in 2 ml of fresh culture medium\. Cell density and viability were determined using a Casy cell counter \(10 µl cell suspension in 10 ml Casyton solution\)\. Neuro\-2a cells were seeded at a density of 20,000 cells per well in 150 µl of complete EMEM into clear, flat\-bottom, white polystyrene 96\-well tissue culture plates \(Corning, 3610\)\. Plates were incubated overnight at 37 °C with 5% CO2to allow for recovery and adhesion\. Following incubation, cytotoxicity was assessed using the CellTox™ Green Cytotoxicity Assay \(Promega\)\. A working medium \(CTG\-EMEM\) was prepared by adding CellTox™ Green Dye \(1000X\) to complete EMEM to a final concentration of 1\.5X\. Spent culture fluid \(100 µl\) was aspirated from the wells and replaced with 100 µl of treatment master mixes prepared in CTG\-EMEM, resulting in a final dye concentration of 1X\. Green fluorescence was quantified using a Cytation 3 imaging plate reader from Biotek/Agilent, with normalization performed between 100% TTX neutralization \(200 µM V \+ 400 µM O, with no TTX\) and 0% TTX neutralization \(200 µM V \+ 400 µM O \+ 200 µM TTX\), following 72 hours of incubation time for osmotic cell lysis to occur\.
Assays utilized a number of combinations of samples and reagents, which were pre\-incubated for 30 minutes prior to exposure to the Neuro\-2a cells \(Supplementary Fig\.[5](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\)\. Controls included CTG\-EMEM\-only \(untreated\) or CTG\-EMEM containing 4% Lysis Solution \(lysis control\), and TTX treatments used 200 nM TTX citrate \(Abcam, ab120055\)\. Wildtype adult hookworm excretory/secretory proteins \(ESPs\) were collected following established protocol[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\. Blood was collected from hamsters infected with iL3s that hatched from wild\-type eggs or transgenic electroporated eggs\. On day 22 post\-infection, hamsters were dissected as described above, and approximately 500\-600 µl of blood was collected via cardiac puncture using a 29 G insulin syringe to obtain a larger volume directly from the heart\. Serum was collected by centrifugation at 4000 ×*g*for 45 minutes at room temperature \(RT\)\. The serum was aliquoted and stored at −20 °C before performing the tetrodotoxin \(TTX\) neutralization using undiluted serum\. We utilized an assay design modified from Chulanetra et al\. 2012[13](https://www.nature.com/articles/s41467-026-73447-9#ref-CR13), in which TTX \(Abcam, Cat \#ab120055; Soluble in water\) activity is quantified by its neutralization of the ion\-channel mediators veratridine \(V\) \(Abcam, Cat \#ab120279; Soluble in ethanol to 100 mM\) and ouabain \(O\) \(Abcam, Cat \#ab120748; Soluble in DMSO to 100 mM, in ethanol to 10 mM, or water to 10 mM\)\. Recombinant s16\-HuScFv was prepared as described above\. All samples were stored at −80 °C when supplied, and working stocks of TTX were prepared fresh daily and stored at 4 °C\. Plates and reagent mixtures were incubated for 30 minutes at 37 °C\. Subsequently, agonist stock solutions were prepared in ethanol: Veratridine \(10 mM; Abcam, ab120279\) and Ouabain octahydrate \(8\.23 mM; Abcam, ab120748\)\. Working solutions were prepared in EMEM \(0\.2 mM Veratridine and 0\.4 mM Ouabain\)\. Cells were treated by adding 50 µl of Veratridine working solution and 50 µl of Ouabain working solution to designated wells\. Plates were sealed and incubated at 37 °C with 5% CO2\. Fluorescence was measured at 1, 6, 24, 48 and 72 hours post\-treatment using a microplate reader with bottom optics\. The excitation filter was set to 485–12 nm and the emission filter to 520 nm, with a gain adjustment between 500 and 700\. Background fluorescence values were subtracted from test samples, and data were plotted as relative fluorescence units \(RFU\)\. For some of the serum samples and matched controls, the Promega CellTox Green Cytotoxicity Assay reagents could not be procured, so the Calcein AM Cell Viability Assay was instead utilized to measure live cell fluorescence following 72 hours of incubation time\.
### Digital image creation
All diagram components were drawn/created in MS PowerPoint\. A hamster image and worm images were captured by the authors during experiments presented in this study, and were used as references for hamster and worm representations\. All other objects were drawn freehand\.
### Mass spectrometry proteomics of hamster serum
One microliter of each infected hamster serum sample was added to 8 M urea and 0\.4 M ammonium bicarbonate buffer, reduced with 4 mM dithiothreitol, and alkylated with 18 mM iodoacetamide\. The solution was then diluted to <2 M urea, and 1 µg of trypsin was added for overnight digestion at 37 °C\. The resulting peptides were desalted using C18 solid\-phase extraction spin columns, and eluates were dried under vacuum using a SpeedVac concentrator\. Dried peptides were reconstituted in 0\.1% formic acid, and concentrations were measured using a NanoDrop assay\. Peptides were loaded onto a Neo trap cartridge coupled to an analytical C18 column \(75 µm × 25 cm, 1\.7 µm; IonOpticks\) and separated using a 35\-minute linear gradient from 2% to 45% solvent B \(0\.1% formic acid in 80% acetonitrile\) on a Vanquish Neo UHPLC system coupled to an Orbitrap Astral mass spectrometer equipped with a FAIMS Pro Duo interface \(Thermo Fisher Scientific, Cat \#FMS03\-10001\)\. Data were processed using Spectronaut 18 \(Biognosys AG\)\. Raw data files were analyzed using directDIA™ against a combined database containing proteins from*Mesocricetus auratus*\(MesAur1\.0 from GenBank[82](https://www.nature.com/articles/s41467-026-73447-9#ref-CR82)\),*A\. ceylanicum*\(genome annotation version described in Uzoechi et al 2023[17](https://www.nature.com/articles/s41467-026-73447-9#ref-CR17)\), and the s16\-HuScFv protein sequence\. Search parameters included trypsin digestion with cleavage after K or R, allowance for up to two missed cleavages, carbamidomethylation of cysteine \(static modification\), and variable modifications including oxidized methionine and Protein N\-terminal acetylation\. Peptide spectrum matches \(PSMs\), peptides, and protein groups were identified using a false discovery rate \(FDR\) threshold of 0\.01, requiring 1 unique peptide per protein\. Differential expression was measured using limma v3\.66\.0[47](https://www.nature.com/articles/s41467-026-73447-9#ref-CR47), with standard settings, and data was sorted, parsed in MS Excel Version 16\.108 \(See Supplementary Data[5](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\)\. Samples sequenced included four biological replicates each of hamster serum from hamsters infected with WT or s16\-HuScFv transgenic hookworms\.
### Statistics & Reproducibility
All samples described as “biological replicates” represent distinct biological samples\. Relative fluorescence values from TTX assay datasets are normally distributed according to a Shapiro\-Wilk test \(W = 0\.9462,*P*= 0\.5828, effect size = 0\.1677; using sample size*n*= 12 biological replicates for the same sample type\), so an ANOVA was used to compare their values\. Significant differences between WT and transgenic samples were tested for qPCR results using two\-tailed*t*\-tests \(equal variance\), using the ∆CT values, before transformation to 2\-∆CTgene expression values\. RNA\-seq differential expression was quantified by DESeq2 FDR\-adjusted*P*≤ 0\.05 and a minimum 2\-fold change in expression\. Source data for all experiments is provided for all datasets in Supplementary Data and Source Data files\. Sample sizes were determined according to the necessary evidence to confirm each individual experiment, while minimizing the number of hamsters that needed to be sacrificed to produce samples\. Replicate counts for RNA\-seq and proteomics analysis were based on standard required biological replicate counts, based on our experience with these datasets\. No data was excluded from the analyses\. The investigators were not blinded to allocation during experiments and outcome assessment\. Blinding was not relevant to this study, as the experimental outcomes relied on objective molecular assays to confirm transgene integration and did not involve subjective evaluations\. Hamsters, worms and eggs were allocated into experimental groups randomly for all experiments\.
### Reporting summary
Further information on research design is available in the[Nature Portfolio Reporting Summary](https://www.nature.com/articles/s41467-026-73447-9#MOESM4)linked to this article\.
## Data availability
The RNA\-seq data generated in this study have been deposited in the NCBI Sequence Read Archive \(SRA\) database under BioProject PRJNA1328670, \[[https://www\.ncbi\.nlm\.nih\.gov/bioproject/PRJNA1328670/](https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1328670/)\]\. The accession information per sample is available in Supplementary Data[4](https://www.nature.com/articles/s41467-026-73447-9#MOESM3), and the processed relative gene expression levels and differential expression statistics for all genes are available in Supplementary Data[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\. The proteomics data generated in this study have been deposited in the iProX repository under accession code[IPX0014753001](https://www.iprox.cn//page/subproject.html?id=IPX0014753001)\. The processed MS proteomics detection level details and a complete list of all hookworm and host proteins detected in each sample are available in Supplementary Data[5](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\. Complete ONT sequencing reads are provided in Supplementary Note[7](https://www.nature.com/articles/s41467-026-73447-9#MOESM1)\. Previously published whole\-worm*A\. ceylanicum*RNA\-seq datasets spanning the life cycle are available from the NCBI Sequence Read Archive \(SRA\) database under BioProject[PRJNA72583](https://www.ncbi.nlm.nih.gov/bioproject/PRJNA72583), accessions numbers SRX1116899\-1116923[14](https://www.nature.com/articles/s41467-026-73447-9#ref-CR14), and the processed relative gene expression levels for all genes across all samples are available in Supplementary Data[1](https://www.nature.com/articles/s41467-026-73447-9#MOESM3)\. The*A\. ceylanicum*genome assembly is available under NCBI BioProject PRJNA231479, assembly accession GCA\_000688135\.1 \[[https://www\.ncbi\.nlm\.nih\.gov/datasets/genome/GCA\_000688135\.1/](https://www.ncbi.nlm.nih.gov/datasets/genome/GCA_000688135.1/)\], and gene annotations were downloaded from WormBase Parasite accession \#PRJNA72583\.WBPS15, annotation version 2020\-04\-WormBase \([https://parasite\.wormbase\.org/Ancylostoma\_ceylanicum\_prjna72583/Info/Index](https://parasite.wormbase.org/Ancylostoma_ceylanicum_prjna72583/Info/Index)\)\. The authors declare that all other data supporting the findings of this study are available within the paper and its supplementary information files\.[Source data](https://www.nature.com/articles/s41467-026-73447-9#Sec43)are provided with this paper\.
## Code availability
The DESeq2 R script for differential gene expression was retrieved from protocols\.io \([https://doi\.org/10\.21203/rs\.3\.pex\-2532/v1](https://doi.org/10.21203/rs.3.pex-2532/v1)\)\. No other custom code was generated or used for any of the analyses described\.
## References
1. Yoon, C\., Ham, Y\. S\., Gil, W\. J\. & Yang, C\. S\. Exploring the potential of*Toxoplasma gondii*in drug development and as a delivery system\.*Exp\. Mol\.Med\.***56**,289–300 \(2024\)\. [Article](https://doi.org/10.1038%2Fs12276-024-01165-7)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2cXisFWrt7g%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=38297164)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC10907749)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Exploring%20the%20potential%20of%20Toxoplasma%20gondii%20in%20drug%20development%20and%20as%20a%20delivery%20system&journal=Exp.%20Mol.Med.&doi=10.1038%2Fs12276-024-01165-7&volume=56&pages=289-300&publication_year=2024&author=Yoon%2CC&author=Ham%2CYS&author=Gil%2CWJ&author=Yang%2CCS)
2. Walther, D\. et al\. Genetically modified helminths as pharmaceutical biofactories\.*Adv\. Parasitol\.***129**, 75–114 \(2025\)\. [Article](https://doi.org/10.1016%2Fbs.apar.2025.08.001)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=41062240)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Genetically%20modified%20helminths%20as%20pharmaceutical%20biofactories&journal=Adv.%20Parasitol.&doi=10.1016%2Fbs.apar.2025.08.001&volume=129&pages=75-114&publication_year=2025&author=Walther%2CD)
3. Beaver, P\. C\. Light, long\-lasting*Necator*infection in a volunteer\.*Am\. J\. Trop\. Med\. Hyg\.***39**, 369–372 \(1988\)\. [Article](https://doi.org/10.4269%2Fajtmh.1988.39.369)[ADS](http://adsabs.harvard.edu/cgi-bin/nph-data_query?link_type=ABSTRACT&bibcode=1988rsgb.conf.....B)[CAS](https://www.nature.com/articles/cas-redirect/1:STN:280:DyaL1M%2FkvVSmsQ%3D%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=3189697)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Light%2C%20long-lasting%20Necator%20infection%20in%20a%20volunteer&journal=Am.%20J.%20Trop.%20Med.%20Hyg.&doi=10.4269%2Fajtmh.1988.39.369&volume=39&pages=369-372&publication_year=1988&author=Beaver%2CPC)
4. Chapman, P\. R\., Giacomin, P\., Loukas, A\. & McCarthy, J\. S\. Experimental human hookworm infection: a narrative historical review\.*PLoS Negl\. Trop\. Dis\.***15**, e0009908 \(2021\)\. [Article](https://doi.org/10.1371%2Fjournal.pntd.0009908)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=34882670)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC8659326)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Experimental%20human%20hookworm%20infection%3A%20a%20narrative%20historical%20review&journal=PLoS%20Negl.%20Trop.%20Dis.&doi=10.1371%2Fjournal.pntd.0009908&volume=15&publication_year=2021&author=Chapman%2CPR&author=Giacomin%2CP&author=Loukas%2CA&author=McCarthy%2CJS)
5. Croese, J\. et al\. Experimental hookworm infection and gluten microchallenge promote tolerance in celiac disease\.*J\. Allergy Clin\. Immunol\.***135**, 508–516 \(2015\)\. [Article](https://doi.org/10.1016%2Fj.jaci.2014.07.022)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2cXhsFyqsrvL)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25248819)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Experimental%20hookworm%20infection%20and%20gluten%20microchallenge%20promote%20tolerance%20in%20celiac%20disease&journal=J.%20Allergy%20Clin.%20Immunol.&doi=10.1016%2Fj.jaci.2014.07.022&volume=135&pages=508-516&publication_year=2015&author=Croese%2CJ)
6. Hoogerwerf, M\. A\. et al\. A randomized controlled trial to investigate safety and variability of egg excretion after repeated controlled human Hookworm infection\.*J\. Infect\. Dis\.***223**, 905–913 \(2021\)\. [Article](https://doi.org/10.1093%2Finfdis%2Fjiaa414)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32645714)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20randomized%20controlled%20trial%20to%20investigate%20safety%20and%20variability%20of%20egg%20excretion%20after%20repeated%20controlled%20human%20Hookworm%20infection&journal=J.%20Infect.%20Dis.&doi=10.1093%2Finfdis%2Fjiaa414&volume=223&pages=905-913&publication_year=2021&author=Hoogerwerf%2CMA)
7. Rosa, B\. A\., McNulty, S\. N\., Mitreva, M\. & Jasmer, D\. P\. Direct experimental manipulation of intestinal cells in Ascaris suum, with minor influences on the global transcriptome\.*Int J\. Parasitol\.***47**, 271–279 \(2017\)\. [Article](https://doi.org/10.1016%2Fj.ijpara.2016.12.005)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2sXjt1KhtL8%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=28223178)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5423655)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Direct%20experimental%20manipulation%20of%20intestinal%20cells%20in%20Ascaris%20suum%2C%20with%20minor%20influences%20on%20the%20global%20transcriptome&journal=Int%20J.%20Parasitol.&doi=10.1016%2Fj.ijpara.2016.12.005&volume=47&pages=271-279&publication_year=2017&author=Rosa%2CBA&author=McNulty%2CSN&author=Mitreva%2CM&author=Jasmer%2CDP)
8. Iqbal, S\., Fosu\-Nyarko, J\. & Jones, M\. G\. K\. Attempt to Silence Genes of the RNAi Pathways of the Root\-Knot Nematode,*Meloidogyne incognita*results in diverse responses including increase and no change in expression of some genes\.*Front\. Plant Sci\.***11**, 328 \(2020\)\. [Article](https://doi.org/10.3389%2Ffpls.2020.00328)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32265973)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7105803)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Attempt%20to%20Silence%20Genes%20of%20the%20RNAi%20Pathways%20of%20the%20Root-Knot%20Nematode%2C%20Meloidogyne%20incognita%20results%20in%20diverse%20responses%20including%20increase%20and%20no%20change%20in%20expression%20of%20some%20genes&journal=Front.%20Plant%20Sci.&doi=10.3389%2Ffpls.2020.00328&volume=11&publication_year=2020&author=Iqbal%2CS&author=Fosu-Nyarko%2CJ&author=Jones%2CMGK)
9. Gang, S\. S\. et al\. Targeted mutagenesis in a human\-parasitic nematode\.*PLoS Pathog\.***13**, e1006675 \(2017\)\. [Article](https://doi.org/10.1371%2Fjournal.ppat.1006675)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=29016680)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5650185)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Targeted%20mutagenesis%20in%20a%20human-parasitic%20nematode&journal=PLoS%20Pathog.&doi=10.1371%2Fjournal.ppat.1006675&volume=13&publication_year=2017&author=Gang%2CSS)
10. Liu, C\., Grote, A\., Ghedin, E\. & Unnasch, T\. R\. CRISPR\-mediated Transfection of*Brugia malayi*\.*PLoS Negl\. Trop\. Dis\.***14**, e0008627 \(2020\)\. [Article](https://doi.org/10.1371%2Fjournal.pntd.0008627)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3cXhvVOhsbfF)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32866158)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7485969)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=CRISPR-mediated%20Transfection%20of%20Brugia%20malayi&journal=PLoS%20Negl.%20Trop.%20Dis.&doi=10.1371%2Fjournal.pntd.0008627&volume=14&publication_year=2020&author=Liu%2CC&author=Grote%2CA&author=Ghedin%2CE&author=Unnasch%2CTR)
11. Sadelain, M\., Papapetrou, E\. P\. & Bushman, F\. D\. Safe harbours for the integration of new DNA in the human genome\.*Nat\. Rev\. Cancer***12**, 51–58 \(2011\)\. [Article](https://doi.org/10.1038%2Fnrc3179)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=22129804)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Safe%20harbours%20for%20the%20integration%20of%20new%20DNA%20in%20the%20human%20genome&journal=Nat.%20Rev.%20Cancer&doi=10.1038%2Fnrc3179&volume=12&pages=51-58&publication_year=2011&author=Sadelain%2CM&author=Papapetrou%2CEP&author=Bushman%2CFD)
12. Koike, A\. & Brindley, P\. J\. CRISPR/Cas genome editing, functional genomics, and diagnostics for parasitic helminths\.*Int J\. Parasitol\.***55**, 741–754 \(2025\)\. [Article](https://doi.org/10.1016%2Fj.ijpara.2025.05.001)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2MXhsVWmu7nF)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=40348052)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC12353640)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=CRISPR%2FCas%20genome%20editing%2C%20functional%20genomics%2C%20and%20diagnostics%20for%20parasitic%20helminths&journal=Int%20J.%20Parasitol.&doi=10.1016%2Fj.ijpara.2025.05.001&volume=55&pages=741-754&publication_year=2025&author=Koike%2CA&author=Brindley%2CPJ)
13. Chulanetra, M\. et al\. Human ScFv that block sodium ion channel activity of tetrodotoxin\.*Toxicon***59**, 272–282 \(2012\)\. [Article](https://doi.org/10.1016%2Fj.toxicon.2011.11.012)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC38XhtVGntrw%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=22126923)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Human%20ScFv%20that%20block%20sodium%20ion%20channel%20activity%20of%20tetrodotoxin&journal=Toxicon&doi=10.1016%2Fj.toxicon.2011.11.012&volume=59&pages=272-282&publication_year=2012&author=Chulanetra%2CM)
14. Bernot, J\. P\. et al\. Transcriptomic analysis of hookworm*Ancylostoma ceylanicum*life cycle stages reveals changes in G\-protein coupled receptor diversity associated with the onset of parasitism\.*Int J\. Parasitol\.***50**, 603–610 \(2020\)\. [Article](https://doi.org/10.1016%2Fj.ijpara.2020.05.003)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3cXhtl2gur3J)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32592811)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7454011)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Transcriptomic%20analysis%20of%20hookworm%20Ancylostoma%20ceylanicum%20life%20cycle%20stages%20reveals%20changes%20in%20G-protein%20coupled%20receptor%20diversity%20associated%20with%20the%20onset%20of%20parasitism&journal=Int%20J.%20Parasitol.&doi=10.1016%2Fj.ijpara.2020.05.003&volume=50&pages=603-610&publication_year=2020&author=Bernot%2CJP)
15. International Helminth Genomes Consortium\. Comparative genomics of the major parasitic worms\.*Nat\. Genet\.***51**, 163–174 \(2019\)\. [Article](https://doi.org/10.1038%2Fs41588-018-0262-1)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1cXitV2qur%2FO)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Comparative%20genomics%20of%20the%20major%20parasitic%20worms&journal=Nat.%20Genet.&doi=10.1038%2Fs41588-018-0262-1&volume=51&pages=163-174&publication_year=2019)
16. Bailey, T\. L\. STREME: Accurate and versatile sequence motif discovery\.*Bioinformatics***37**, 2834–2840 \(2021\)\. [Article](https://doi.org/10.1093%2Fbioinformatics%2Fbtab203)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB38Xitl2lsr8%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=33760053)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC8479671)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=STREME%3A%20Accurate%20and%20versatile%20sequence%20motif%20discovery&journal=Bioinformatics&doi=10.1093%2Fbioinformatics%2Fbtab203&volume=37&pages=2834-2840&publication_year=2021&author=Bailey%2CTL)
17. Uzoechi, S\. C\. et al\. Excretory/Secretory Proteome of Females and Males of the Hookworm Ancylostoma ceylanicum\.*Pathogens***12**, 95 \(2023\)\. [Article](https://doi.org/10.3390%2Fpathogens12010095)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3sXjtVOrsr4%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=36678443)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC9865600)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Excretory%2FSecretory%20Proteome%20of%20Females%20and%20Males%20of%20the%20Hookworm%20Ancylostoma%20ceylanicum&journal=Pathogens&doi=10.3390%2Fpathogens12010095&volume=12&publication_year=2023&author=Uzoechi%2CSC)
18. Gupta, S\., Stamatoyannopoulos, J\. A\., Bailey, T\. L\. & Noble, W\. S\. Quantifying similarity between motifs\.*Genome Biol\.***8**, R24 \(2007\)\. [Article](https://link.springer.com/doi/10.1186/gb-2007-8-2-r24)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=17324271)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1852410)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Quantifying%20similarity%20between%20motifs&journal=Genome%20Biol.&doi=10.1186%2Fgb-2007-8-2-r24&volume=8&publication_year=2007&author=Gupta%2CS&author=Stamatoyannopoulos%2CJA&author=Bailey%2CTL&author=Noble%2CWS)
19. Almagro Armenteros, J\. J\. et al\. SignalP 5\.0 improves signal peptide predictions using deep neural networks\.*Nat\. Biotechnol\.***37**, 420–423 \(2019\)\. [Article](https://doi.org/10.1038%2Fs41587-019-0036-z)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1MXosV2qs78%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30778233)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=SignalP%205.0%20improves%20signal%20peptide%20predictions%20using%20deep%20neural%20networks&journal=Nat.%20Biotechnol.&doi=10.1038%2Fs41587-019-0036-z&volume=37&pages=420-423&publication_year=2019&author=Almagro%20Armenteros%2CJJ)
20. Logan, J\. et al\. Comprehensive analysis of the secreted proteome of adult*Necator americanus*hookworms\.*PLoS Negl\. Trop\. Dis\.***14**, e0008237 \(2020\)\. [Article](https://doi.org/10.1371%2Fjournal.pntd.0008237)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3cXit1GlsbvM)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32453752)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7274458)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Comprehensive%20analysis%20of%20the%20secreted%20proteome%20of%20adult%20Necator%20americanus%20hookworms&journal=PLoS%20Negl.%20Trop.%20Dis.&doi=10.1371%2Fjournal.pntd.0008237&volume=14&publication_year=2020&author=Logan%2CJ)
21. Morante, T\., Shepherd, C\., Constantinoiu, C\., Loukas, A\. & Sotillo, J\. Revisiting the*Ancylostoma Caninum*Secretome Provides New Information on Hookworm\-Host Interactions\.*Proteomics***17**, 1700186 \(2017\)\. [Article](https://doi.org/10.1002%2Fpmic.201700186)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Revisiting%20the%20Ancylostoma%20Caninum%20Secretome%20Provides%20New%20Information%20on%20Hookworm-Host%20Interactions&journal=Proteomics&doi=10.1002%2Fpmic.201700186&volume=17&publication_year=2017&author=Morante%2CT&author=Shepherd%2CC&author=Constantinoiu%2CC&author=Loukas%2CA&author=Sotillo%2CJ)
22. Miele, A\. E\. et al\. A comparative analysis of secreted protein disulfide isomerases from the tropical co\-endemic parasites*Schistosoma mansoni*and*Leishmania major*\.*Sci\. Rep\.***9**, 9568 \(2019\)\. [Article](https://doi.org/10.1038%2Fs41598-019-45709-8)[ADS](http://adsabs.harvard.edu/cgi-bin/nph-data_query?link_type=ABSTRACT&bibcode=2019NatSR...9.9568M)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31267027)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6606611)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20comparative%20analysis%20of%20secreted%20protein%20disulfide%20isomerases%20from%20the%20tropical%20co-endemic%20parasites%20Schistosoma%20mansoni%20and%20Leishmania%20major&journal=Sci.%20Rep.&doi=10.1038%2Fs41598-019-45709-8&volume=9&publication_year=2019&author=Miele%2CAE)
23. Fonseca, C\. T\., Braz Figueiredo Carvalho, G\., Carvalho Alves, C\. & de Melo, T\. T\.*Schistosoma*tegument proteins in vaccine and diagnosis development: an update\.*J\. Parasitol\. Res***2012**, 541268 \(2012\)\. [Article](https://doi.org/10.1155%2F2012%2F541268)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=23125917)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3483795)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Schistosoma%20tegument%20proteins%20in%20vaccine%20and%20diagnosis%20development%3A%20an%20update&journal=J.%20Parasitol.%20Res&doi=10.1155%2F2012%2F541268&volume=2012&publication_year=2012&author=Fonseca%2CCT&author=Braz%20Figueiredo%20Carvalho%2CG&author=Carvalho%20Alves%2CC&author=Melo%2CTT)
24. Tian, Z\. L\., Wang, Z\. H\., Maria, M\., Qu, N\. & Zheng, J\. W\.*Meloidogyne graminicola*protein disulfide isomerase may be a nematode effector and is involved in protection against oxidative damage\.*Sci\. Rep\.***9**, 11949 \(2019\)\. [Article](https://doi.org/10.1038%2Fs41598-019-48474-w)[ADS](http://adsabs.harvard.edu/cgi-bin/nph-data_query?link_type=ABSTRACT&bibcode=2019NatSR...911949T)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31420562)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6697734)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Meloidogyne%20graminicola%20protein%20disulfide%20isomerase%20may%20be%20a%20nematode%20effector%20and%20is%20involved%20in%20protection%20against%20oxidative%20damage&journal=Sci.%20Rep.&doi=10.1038%2Fs41598-019-48474-w&volume=9&publication_year=2019&author=Tian%2CZL&author=Wang%2CZH&author=Maria%2CM&author=Qu%2CN&author=Zheng%2CJW)
25. Castro\-Mondragon, J\. A\. et al\. JASPAR 2022: the 9th release of the open\-access database of transcription factor binding profiles\.*Nucleic Acids Res\.***50**, D165–D173 \(2022\)\. [Article](https://doi.org/10.1093%2Fnar%2Fgkab1113)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB38Xis1eisbc%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=34850907)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC8728201)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=JASPAR%202022%3A%20the%209th%20release%20of%20the%20open-access%20database%20of%20transcription%20factor%20binding%20profiles&journal=Nucleic%20Acids%20Res.&doi=10.1093%2Fnar%2Fgkab1113&volume=50&pages=D165-D173&publication_year=2022&author=Castro-Mondragon%2CJA)
26. Wong, Y\. et al\. Proteomic characterization and comparison of the infective and adult life stage secretomes from*Necator americanus*and*Ancylostoma ceylanicum*\.*PLoS Negl\. Trop\. Dis\.***19**, e0012780 \(2025\)\. [Article](https://doi.org/10.1371%2Fjournal.pntd.0012780)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2MXoslCmu74%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=39832284)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC11745416)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Proteomic%20characterization%20and%20comparison%20of%20the%20infective%20and%20adult%20life%20stage%20secretomes%20from%20Necator%20americanus%20and%20Ancylostoma%20ceylanicum&journal=PLoS%20Negl.%20Trop.%20Dis.&doi=10.1371%2Fjournal.pntd.0012780&volume=19&publication_year=2025&author=Wong%2CY)
27. Labun, K\., Krause, M\., Torres Cleuren, Y\. & Valen, E\. CRISPR Genome Editing Made Easy Through the CHOPCHOP Website\.*Curr\. Protoc\.***1**, e46 \(2021\)\. [Article](https://doi.org/10.1002%2Fcpz1.46)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3MXpsFGht78%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=33905612)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=CRISPR%20Genome%20Editing%20Made%20Easy%20Through%20the%20CHOPCHOP%20Website&journal=Curr.%20Protoc.&doi=10.1002%2Fcpz1.46&volume=1&publication_year=2021&author=Labun%2CK&author=Krause%2CM&author=Torres%20Cleuren%2CY&author=Valen%2CE)
28. Ittiprasert, W\. et al\. Targeted insertion and reporter transgene activity at a gene safe harbor of the human blood fluke,*Schistosoma mansoni*\.*Cell Rep\. Methods***3**, 100535 \(2023\)\. [Article](https://doi.org/10.1016%2Fj.crmeth.2023.100535)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=37533651)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC10391569)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Targeted%20insertion%20and%20reporter%20transgene%20activity%20at%20a%20gene%20safe%20harbor%20of%20the%20human%20blood%20fluke%2C%20Schistosoma%20mansoni&journal=Cell%20Rep.%20Methods&doi=10.1016%2Fj.crmeth.2023.100535&volume=3&publication_year=2023&author=Ittiprasert%2CW)
29. Ittiprasert, W\., Moescheid, M\. M\., Mann, V\. H\. & Brindley, P\. J\. Multiplexed CRISPR\-Cas9 protocol for large transgene integration into the*Schistosoma mansoni*genome\.*STAR Protoc\.***5**, 102886 \(2024\)\. [Article](https://doi.org/10.1016%2Fj.xpro.2024.102886)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2cXktFWjt7o%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=38354082)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC10876972)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Multiplexed%20CRISPR-Cas9%20protocol%20for%20large%20transgene%20integration%20into%20the%20Schistosoma%20mansoni%20genome&journal=STAR%20Protoc.&doi=10.1016%2Fj.xpro.2024.102886&volume=5&publication_year=2024&author=Ittiprasert%2CW&author=Moescheid%2CMM&author=Mann%2CVH&author=Brindley%2CPJ)
30. Yuan, B\. et al\. Modulation of the microhomology\-mediated end joining pathway suppresses large deletions and enhances homology\-directed repair following CRISPR\-Cas9\-induced DNA breaks\.*BMC Biol\.***22**, 101 \(2024\)\. [Article](https://link.springer.com/doi/10.1186/s12915-024-01896-z)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2cXhtVWrt7fM)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=38685010)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC11059712)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Modulation%20of%20the%20microhomology-mediated%20end%20joining%20pathway%20suppresses%20large%20deletions%20and%20enhances%20homology-directed%20repair%20following%20CRISPR-Cas9-induced%20DNA%20breaks&journal=BMC%20Biol.&doi=10.1186%2Fs12915-024-01896-z&volume=22&publication_year=2024&author=Yuan%2CB)
31. Liu, C\. et al\. Development of a toolkit for piggyBac\-mediated integrative transfection of the human filarial parasite*Brugia malayi*\.*PLoS Negl\. Trop\. Dis\.***12**, e0006509 \(2018\)\. [Article](https://doi.org/10.1371%2Fjournal.pntd.0006509)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=29782496)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5983866)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Development%20of%20a%20toolkit%20for%20piggyBac-mediated%20integrative%20transfection%20of%20the%20human%20filarial%20parasite%20Brugia%20malayi&journal=PLoS%20Negl.%20Trop.%20Dis.&doi=10.1371%2Fjournal.pntd.0006509&volume=12&publication_year=2018&author=Liu%2CC)
32. Hawdon, J\. M\., Narasimhan, S\. & Hotez, P\. J\. Ancylostoma secreted protein 2: cloning and characterization of a second member of a family of nematode secreted proteins from*Ancylostoma caninum*\.*Mol\. Biochem\. Parasitol\.***99**, 149–165 \(1999\)\. [Article](https://doi.org/10.1016%2FS0166-6851%2899%2900011-0)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DyaK1MXjt1Snsb4%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=10340481)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Ancylostoma%20secreted%20protein%202%3A%20cloning%20and%20characterization%20of%20a%20second%20member%20of%20a%20family%20of%20nematode%20secreted%20proteins%20from%20Ancylostoma%20caninum&journal=Mol.%20Biochem.%20Parasitol.&doi=10.1016%2FS0166-6851%2899%2900011-0&volume=99&pages=149-165&publication_year=1999&author=Hawdon%2CJM&author=Narasimhan%2CS&author=Hotez%2CPJ)
33. El Atab, O\. et al\.*Necator americanus*Ancylostoma Secreted Protein\-2 \(Na\-ASP\-2\) Binds an Ascaroside \(ascr\#3\) in Its Fatty Acid Binding Site\.*Front\. Chem\.***8**, 608296 \(2020\)\. [Article](https://doi.org/10.3389%2Ffchem.2020.608296)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3MXovVemtr4%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=33392151)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7773830)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Necator%20americanus%20Ancylostoma%20Secreted%20Protein-2%20%28Na-ASP-2%29%20Binds%20an%20Ascaroside%20%28ascr%233%29%20in%20Its%20Fatty%20Acid%20Binding%20Site&journal=Front.%20Chem.&doi=10.3389%2Ffchem.2020.608296&volume=8&publication_year=2020&author=El%20Atab%2CO)
34. Wei, J\. et al\. The hookworm*Ancylostoma ceylanicum*intestinal transcriptome provides a platform for selecting drug and vaccine candidates\.*Parasit\. Vectors***9**, 518 \(2016\)\. [Article](https://link.springer.com/doi/10.1186/s13071-016-1795-8)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=27677574)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5039805)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=The%20hookworm%20Ancylostoma%20ceylanicum%20intestinal%20transcriptome%20provides%20a%20platform%20for%20selecting%20drug%20and%20vaccine%20candidates&journal=Parasit.%20Vectors&doi=10.1186%2Fs13071-016-1795-8&volume=9&publication_year=2016&author=Wei%2CJ)
35. Mieszczanek, J\., Harrison, L\. M\. & Cappello, M\.*Ancylostoma ceylanicum*anticoagulant peptide\-1: role of the predicted reactive site amino acid in mediating inhibition of coagulation factors Xa and VIIa\.*Mol\. Biochem\. Parasitol\.***137**, 151–159 \(2004\)\. [Article](https://doi.org/10.1016%2Fj.molbiopara.2004.05.011)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD2cXmtVWju7Y%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=15279961)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Ancylostoma%20ceylanicum%20anticoagulant%20peptide-1%3A%20role%20of%20the%20predicted%20reactive%20site%20amino%20acid%20in%20mediating%20inhibition%20of%20coagulation%20factors%20Xa%20and%20VIIa&journal=Mol.%20Biochem.%20Parasitol.&doi=10.1016%2Fj.molbiopara.2004.05.011&volume=137&pages=151-159&publication_year=2004&author=Mieszczanek%2CJ&author=Harrison%2CLM&author=Cappello%2CM)
36. Verma, M\. et al\. A short translational ramp determines the efficiency of protein synthesis\.*Nat\. Commun\.***10**, 5774 \(2019\)\. [Article](https://doi.org/10.1038%2Fs41467-019-13810-1)[ADS](http://adsabs.harvard.edu/cgi-bin/nph-data_query?link_type=ABSTRACT&bibcode=2019NatCo..10.5774V)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1MXisVKgsb7E)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31852903)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6920384)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20short%20translational%20ramp%20determines%20the%20efficiency%20of%20protein%20synthesis&journal=Nat.%20Commun.&doi=10.1038%2Fs41467-019-13810-1&volume=10&publication_year=2019&author=Verma%2CM)
37. Wibisono, P\., Liu, Y\. & Sun, J\. A novel in vitro*Caenorhabditis elegans*transcription system\.*BMC Mol\. Cell Biol\.***21**, 87 \(2020\)\. [Article](https://link.springer.com/doi/10.1186/s12860-020-00332-8)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3MXksFyrtw%3D%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=33256604)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7706227)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20novel%20in%20vitro%20Caenorhabditis%20elegans%20transcription%20system&journal=BMC%20Mol.%20Cell%20Biol.&doi=10.1186%2Fs12860-020-00332-8&volume=21&publication_year=2020&author=Wibisono%2CP&author=Liu%2CY&author=Sun%2CJ)
38. Moguel, B\. et al\. Transient transgenesis of the tapeworm*Taenia crassiceps*\.*Springerplus***4**, 496 \(2015\)\. [Article](https://link.springer.com/doi/10.1186/s40064-015-1278-y)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=26389021)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4571025)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Transient%20transgenesis%20of%20the%20tapeworm%20Taenia%20crassiceps&journal=Springerplus&doi=10.1186%2Fs40064-015-1278-y&volume=4&publication_year=2015&author=Moguel%2CB)
39. Liang, S\., Knight, M\. & Jolly, E\. R\. Polyethyleneimine mediated DNA transfection in schistosome parasites and regulation of the WNT signaling pathway by a dominant\-negative SmMef2\.*PLoS Negl\. Trop\. Dis\.***7**, e2332 \(2013\)\. [Article](https://doi.org/10.1371%2Fjournal.pntd.0002332)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3sXht1CmtL7N)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=23936566)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3723562)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Polyethyleneimine%20mediated%20DNA%20transfection%20in%20schistosome%20parasites%20and%20regulation%20of%20the%20WNT%20signaling%20pathway%20by%20a%20dominant-negative%20SmMef2&journal=PLoS%20Negl.%20Trop.%20Dis.&doi=10.1371%2Fjournal.pntd.0002332&volume=7&publication_year=2013&author=Liang%2CS&author=Knight%2CM&author=Jolly%2CER)
40. Liu, Y\. et al\. Biallelic insertion of a transcriptional terminator via the CRISPR/Cas9 system efficiently silences expression of protein\-coding and non\-coding RNA genes\.*J\. Biol\. Chem\.***292**, 5624–5633 \(2017\)\. [Article](https://doi.org/10.1074%2Fjbc.M116.769034)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2sXlslCku78%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=28196870)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5392558)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Biallelic%20insertion%20of%20a%20transcriptional%20terminator%20via%20the%20CRISPR%2FCas9%20system%20efficiently%20silences%20expression%20of%20protein-coding%20and%20non-coding%20RNA%20genes&journal=J.%20Biol.%20Chem.&doi=10.1074%2Fjbc.M116.769034&volume=292&pages=5624-5633&publication_year=2017&author=Liu%2CY)
41. Brees, C\. & Fransen, M\. A cost\-effective approach to microporate mammalian cells with the Neon Transfection System\.*Anal\. Biochem\.***466**, 49–50 \(2014\)\. [Article](https://doi.org/10.1016%2Fj.ab.2014.08.017)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2cXhsFCns7fK)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25172131)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20cost-effective%20approach%20to%20microporate%20mammalian%20cells%20with%20the%20Neon%20Transfection%20System&journal=Anal.%20Biochem.&doi=10.1016%2Fj.ab.2014.08.017&volume=466&pages=49-50&publication_year=2014&author=Brees%2CC&author=Fransen%2CM)
42. Anders, S\. & Huber, W\. Differential expression analysis for sequence count data\.*Genome Biol\.***11**, R106 \(2010\)\. [Article](https://link.springer.com/doi/10.1186/gb-2010-11-10-r106)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3cXhsVahs7bI)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=20979621)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3218662)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Differential%20expression%20analysis%20for%20sequence%20count%20data&journal=Genome%20Biol.&doi=10.1186%2Fgb-2010-11-10-r106&volume=11&publication_year=2010&author=Anders%2CS&author=Huber%2CW)
43. Chavez, V\., Mohri\-Shiomi, A\. & Garsin, D\. A\. Ce\-Duox1/BLI\-3 generates reactive oxygen species as a protective innate immune mechanism in*Caenorhabditis elegans*\.*Infect\. Immun\.***77**, 4983–4989 \(2009\)\. [Article](https://doi.org/10.1128%2FIAI.00627-09)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD1MXhtl2itrnL)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=19687201)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2772517)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Ce-Duox1%2FBLI-3%20generates%20reactive%20oxygen%20species%20as%20a%20protective%20innate%20immune%20mechanism%20in%20Caenorhabditis%20elegans&journal=Infect.%20Immun.&doi=10.1128%2FIAI.00627-09&volume=77&pages=4983-4989&publication_year=2009&author=Chavez%2CV&author=Mohri-Shiomi%2CA&author=Garsin%2CDA)
44. de Voer, G\. et al\. Deletion of the*Caenorhabditis elegans*homologues of the CLN3 gene, involved in human juvenile neuronal ceroid lipofuscinosis, causes a mild progeric phenotype\.*J\. Inherit\. Metab\. Dis\.***28**, 1065–1080 \(2005\)\. [Article](https://link.springer.com/doi/10.1007/s10545-005-0125-7)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD28Xnt1GhsQ%3D%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=16435200)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Deletion%20of%20the%20Caenorhabditis%20elegans%20homologues%20of%20the%20CLN3%20gene%2C%20involved%20in%20human%20juvenile%20neuronal%20ceroid%20lipofuscinosis%2C%20causes%20a%20mild%20progeric%20phenotype&journal=J.%20Inherit.%20Metab.%20Dis.&doi=10.1007%2Fs10545-005-0125-7&volume=28&pages=1065-1080&publication_year=2005&author=Voer%2CG)
45. Bruderer, R\. et al\. Extending the limits of quantitative proteome profiling with data\-independent acquisition and application to acetaminophen\-treated three\-dimensional liver microtissues\.*Mol\. Cell Proteom\.***14**, 1400–1410 \(2015\)\. [Article](https://doi.org/10.1074%2Fmcp.M114.044305)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2MXotVWjsbo%3D)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Extending%20the%20limits%20of%20quantitative%20proteome%20profiling%20with%20data-independent%20acquisition%20and%20application%20to%20acetaminophen-treated%20three-dimensional%20liver%20microtissues&journal=Mol.%20Cell%20Proteom.&doi=10.1074%2Fmcp.M114.044305&volume=14&pages=1400-1410&publication_year=2015&author=Bruderer%2CR)
46. Wen, B\. et al\. Assessment of false discovery rate control in tandem mass spectrometry analysis using entrapment\.*Nat\. Methods***22**, 1454–1463 \(2025\)\. [Article](https://doi.org/10.1038%2Fs41592-025-02719-x)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2MXhs1ansrvK)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=40524023)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC12240826)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Assessment%20of%20false%20discovery%20rate%20control%20in%20tandem%20mass%20spectrometry%20analysis%20using%20entrapment&journal=Nat.%20Methods&doi=10.1038%2Fs41592-025-02719-x&volume=22&pages=1454-1463&publication_year=2025&author=Wen%2CB)
47. Ritchie, M\. E\. et al\. limma powers differential expression analyses for RNA\-sequencing and microarray studies\.*Nucleic Acids Res\.***43**, e47 \(2015\)\. [Article](https://doi.org/10.1093%2Fnar%2Fgkv007)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25605792)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4402510)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=limma%20powers%20differential%20expression%20analyses%20for%20RNA-sequencing%20and%20microarray%20studies&journal=Nucleic%20Acids%20Res.&doi=10.1093%2Fnar%2Fgkv007&volume=43&publication_year=2015&author=Ritchie%2CME)
48. Wang, T\. et al\. The Proteome and Lipidome of Extracellular Vesicles from*Haemonchus contortus*to Underpin Explorations of Host\-Parasite Cross\-Talk\.*Int J\. Mol\. Sci\.***24**, 10955 \(2023\)\. [Article](https://doi.org/10.3390%2Fijms241310955)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3sXhsFWmsLjO)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=37446130)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC10341815)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=The%20Proteome%20and%20Lipidome%20of%20Extracellular%20Vesicles%20from%20Haemonchus%20contortus%20to%20Underpin%20Explorations%20of%20Host-Parasite%20Cross-Talk&journal=Int%20J.%20Mol.%20Sci.&doi=10.3390%2Fijms241310955&volume=24&publication_year=2023&author=Wang%2CT)
49. Eichenberger, R\. M\. et al\. Hookworm Secreted Extracellular Vesicles Interact With Host Cells and Prevent Inducible Colitis in Mice\.*Front Immunol\.***9**, 850 \(2018\)\. [Article](https://doi.org/10.3389%2Ffimmu.2018.00850)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=29760697)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5936971)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Hookworm%20Secreted%20Extracellular%20Vesicles%20Interact%20With%20Host%20Cells%20and%20Prevent%20Inducible%20Colitis%20in%20Mice&journal=Front%20Immunol.&doi=10.3389%2Ffimmu.2018.00850&volume=9&publication_year=2018&author=Eichenberger%2CRM)
50. Ancarola, M\. E\. et al\. A comparative analysis of the protein cargo of extracellular vesicles from Helminth parasites\.*Life***13**, 2286 \(2023\)\. [Article](https://doi.org/10.3390%2Flife13122286)[ADS](http://adsabs.harvard.edu/cgi-bin/nph-data_query?link_type=ABSTRACT&bibcode=2023Life...13.2286A)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2cXhtlKksbc%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=38137887)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC10744797)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20comparative%20analysis%20of%20the%20protein%20cargo%20of%20extracellular%20vesicles%20from%20Helminth%20parasites&journal=Life&doi=10.3390%2Flife13122286&volume=13&publication_year=2023&author=Ancarola%2CME)
51. Narahashi, T\., Moore, J\. W\. & Scott, W\. R\. Tetrodotoxin blockage of sodium conductance increase in lobster Giant Axons\.*J\. Gen\. Physiol\.***47**, 965–974 \(1964\)\. [Article](https://doi.org/10.1085%2Fjgp.47.5.965)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DyaF2cXkt1Okt7s%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=14155438)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2195365)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Tetrodotoxin%20blockage%20of%20sodium%20conductance%20increase%20in%20lobster%20Giant%20Axons&journal=J.%20Gen.%20Physiol.&doi=10.1085%2Fjgp.47.5.965&volume=47&pages=965-974&publication_year=1964&author=Narahashi%2CT&author=Moore%2CJW&author=Scott%2CWR)
52. Noguchi, T\. & Ebesu, J\. S\. M\. Puffer poisoning: epidemiology and treatment\.*J\. Toxicol\.: Toxin Rev\.***20**, 1–10 \(2001\)\. [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD3MXkt1Gntb4%3D)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Puffer%20poisoning%3A%20epidemiology%20and%20treatment&journal=J.%20Toxicol.%3A%20Toxin%20Rev.&volume=20&pages=1-10&publication_year=2001&author=Noguchi%2CT&author=Ebesu%2CJSM)
53. Bharti, S\., Rosa, B\. A\., Martin, J\. & Mitreva, M\. Single\-cell transcriptome atlas of adult male and female human hookworm*Ancylostoma ceylanicum*\.*iScience***28**, 113846 \(2025\)\. [Article](https://doi.org/10.1016%2Fj.isci.2025.113846)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2MXjt1GisLzK)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=41362773)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC12682278)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Single-cell%20transcriptome%20atlas%20of%20adult%20male%20and%20female%20human%20hookworm%20Ancylostoma%20ceylanicum&journal=iScience&doi=10.1016%2Fj.isci.2025.113846&volume=28&publication_year=2025&author=Bharti%2CS&author=Rosa%2CBA&author=Martin%2CJ&author=Mitreva%2CM)
54. Durette\-Desset, M\. C\., Beveridge, I\. & Spratt, D\. M\. The origins and evolutionary expansion of the Strongylida \(Nematoda\)\.*Int J\. Parasitol\.***24**, 1139–1165 \(1994\)\. [Article](https://doi.org/10.1016%2F0020-7519%2894%2990188-0)[CAS](https://www.nature.com/articles/cas-redirect/1:STN:280:DyaK2M3kslemuw%3D%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=7729974)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=The%20origins%20and%20evolutionary%20expansion%20of%20the%20Strongylida%20%28Nematoda%29&journal=Int%20J.%20Parasitol.&doi=10.1016%2F0020-7519%2894%2990188-0&volume=24&pages=1139-1165&publication_year=1994&author=Durette-Desset%2CMC&author=Beveridge%2CI&author=Spratt%2CDM)
55. Giacomin, P\., Croese, J\., Krause, L\., Loukas, A\. & Cantacessi, C\. Suppression of inflammation by helminths: a role for the gut microbiota?*Philos\. Trans\. R\. Soc\. Lond\. B Biol\. Sci\.***370**, 20140296 \(2015\)\. [Article](https://doi.org/10.1098%2Frstb.2014.0296)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=26150662)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4528494)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Suppression%20of%20inflammation%20by%20helminths%3A%20a%20role%20for%20the%20gut%20microbiota%3F&journal=Philos.%20Trans.%20R.%20Soc.%20Lond.%20B%20Biol.%20Sci.&doi=10.1098%2Frstb.2014.0296&volume=370&publication_year=2015&author=Giacomin%2CP&author=Croese%2CJ&author=Krause%2CL&author=Loukas%2CA&author=Cantacessi%2CC)
56. Mules, T\. C\. et al\. Controlled Hookworm infection for medication\-free maintenance in patients with ulcerative colitis: a pilot, double\-blind, randomized control trial\.*Inflamm\. Bowel Dis\.***30**, 735–745 \(2024\)\. [Article](https://doi.org/10.1093%2Fibd%2Fizad110)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=37318363)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC11063543)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Controlled%20Hookworm%20infection%20for%20medication-free%20maintenance%20in%20patients%20with%20ulcerative%20colitis%3A%20a%20pilot%2C%20double-blind%2C%20randomized%20control%20trial&journal=Inflamm.%20Bowel%20Dis.&doi=10.1093%2Fibd%2Fizad110&volume=30&pages=735-745&publication_year=2024&author=Mules%2CTC)
57. Cantacessi, C\. et al\. Impact of experimental hookworm infection on the human gut microbiota\.*J\. Infect\. Dis\.***210**, 1431–1434 \(2014\)\. [Article](https://doi.org/10.1093%2Finfdis%2Fjiu256)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC28XitVyhtLzP)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=24795483)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4195438)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Impact%20of%20experimental%20hookworm%20infection%20on%20the%20human%20gut%20microbiota&journal=J.%20Infect.%20Dis.&doi=10.1093%2Finfdis%2Fjiu256&volume=210&pages=1431-1434&publication_year=2014&author=Cantacessi%2CC)
58. Ramchandani, R\., Lucyshyn, R\., Linton, S\. & Ellis, A\. K\. Breaking the mold: nontraditional approaches to allergen immunotherapy for environmental allergens\.*Immunotherapy***16**, 1153–1169 \(2024\)\. [Article](https://doi.org/10.1080%2F1750743X.2024.2408216)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2MXhs1Gqu7vP)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=39382452)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC11633400)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Breaking%20the%20mold%3A%20nontraditional%20approaches%20to%20allergen%20immunotherapy%20for%20environmental%20allergens&journal=Immunotherapy&doi=10.1080%2F1750743X.2024.2408216&volume=16&pages=1153-1169&publication_year=2024&author=Ramchandani%2CR&author=Lucyshyn%2CR&author=Linton%2CS&author=Ellis%2CAK)
59. Singh, R\. Analysis of synonymous codon usage bias in*Ancylostoma ceylanicum*\.*Gene Rep\.***24**, 101290 \(2021\)\. [Article](https://doi.org/10.1016%2Fj.genrep.2021.101290)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB38XhtlWltbfN)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Analysis%20of%20synonymous%20codon%20usage%20bias%20in%20Ancylostoma%20ceylanicum&journal=Gene%20Rep.&doi=10.1016%2Fj.genrep.2021.101290&volume=24&publication_year=2021&author=Singh%2CR)
60. Jourdan, P\. M\., Lamberton, P\. H\. L\., Fenwick, A\. & Addiss, D\. G\. Soil\-transmitted helminth infections\.*Lancet***391**, 252–265 \(2018\)\. [Article](https://doi.org/10.1016%2FS0140-6736%2817%2931930-X)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=28882382)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Soil-transmitted%20helminth%20infections&journal=Lancet&doi=10.1016%2FS0140-6736%2817%2931930-X&volume=391&pages=252-265&publication_year=2018&author=Jourdan%2CPM&author=Lamberton%2CPHL&author=Fenwick%2CA&author=Addiss%2CDG)
61. Lok, J\. B\. CRISPR/Cas9 mutagenesis and expression of dominant mutant transgenes as functional genomic approaches in parasitic nematodes\.*Front\. Genet\.***10**, 656 \(2019\)\. [Article](https://doi.org/10.3389%2Ffgene.2019.00656)[ADS](http://adsabs.harvard.edu/cgi-bin/nph-data_query?link_type=ABSTRACT&bibcode=2019FrCh....7..656L)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3cXhsFGhsr0%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31379923)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6646703)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=CRISPR%2FCas9%20mutagenesis%20and%20expression%20of%20dominant%20mutant%20transgenes%20as%20functional%20genomic%20approaches%20in%20parasitic%20nematodes&journal=Front.%20Genet.&doi=10.3389%2Ffgene.2019.00656&volume=10&publication_year=2019&author=Lok%2CJB)
62. Patel, R\. et al\. The generation of stable transgenic lines in the human\-infective nematode*Strongyloides stercoralis*\.*G3***14**, jkae122 \(2024\)\. [Article](https://doi.org/10.1093%2Fg3journal%2Fjkae122)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2MXht1Wms73N)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=38839055)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC11304987)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=The%20generation%20of%20stable%20transgenic%20lines%20in%20the%20human-infective%20nematode%20Strongyloides%20stercoralis&journal=G3&doi=10.1093%2Fg3journal%2Fjkae122&volume=14&publication_year=2024&author=Patel%2CR)
63. Vacca, F\. et al\. Controlled infection with cryopreserved human hookworm induces CTLA\-4 expression on Tregs and upregulates tryptophan metabolism\.*Gut Microbes***16**, 2416517 \(2024\)\. [Article](https://doi.org/10.1080%2F19490976.2024.2416517)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=39411786)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC11485773)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Controlled%20infection%20with%20cryopreserved%20human%20hookworm%20induces%20CTLA-4%20expression%20on%20Tregs%20and%20upregulates%20tryptophan%20metabolism&journal=Gut%20Microbes&doi=10.1080%2F19490976.2024.2416517&volume=16&publication_year=2024&author=Vacca%2CF)
64. Schwarz, E\. M\. et al\. The genome and transcriptome of the zoonotic hookworm*Ancylostoma ceylanicum*identify infection\-specific gene families\.*Nat\. Genet\.***47**, 416–422 \(2015\)\. [Article](https://doi.org/10.1038%2Fng.3237)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2MXjs1Gqs7s%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25730766)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4617383)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=The%20genome%20and%20transcriptome%20of%20the%20zoonotic%20hookworm%20Ancylostoma%20ceylanicum%20identify%20infection-specific%20gene%20families&journal=Nat.%20Genet.&doi=10.1038%2Fng.3237&volume=47&pages=416-422&publication_year=2015&author=Schwarz%2CEM)
65. Magrini, V\. et al\. Improving eukaryotic genome annotation using single\-molecule mRNA sequencing\.*BMC Genomics***19**, 172 \(2018\)\. [Article](https://link.springer.com/doi/10.1186/s12864-018-4555-7)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=29495964)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5833154)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Improving%20eukaryotic%20genome%20annotation%20using%20single-molecule%20mRNA%20sequencing&journal=BMC%20Genomics&doi=10.1186%2Fs12864-018-4555-7&volume=19&publication_year=2018&author=Magrini%2CV)
66. Howe, K\. L\., Bolt, B\. J\., Shafie, M\., Kersey, P\. & Berriman, M\. WormBase ParaSite \- a comprehensive resource for helminth genomics\.*Mol\. Biochem\. Parasitol\.***215**, 2–10 \(2017\)\. [Article](https://doi.org/10.1016%2Fj.molbiopara.2016.11.005)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC28XitVSmsb%2FF)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=27899279)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=WormBase%20ParaSite%20-%20a%20comprehensive%20resource%20for%20helminth%20genomics&journal=Mol.%20Biochem.%20Parasitol.&doi=10.1016%2Fj.molbiopara.2016.11.005&volume=215&pages=2-10&publication_year=2017&author=Howe%2CKL&author=Bolt%2CBJ&author=Shafie%2CM&author=Kersey%2CP&author=Berriman%2CM)
67. Bolger, A\. M\., Lohse, M\. & Usadel, B\. Trimmomatic: a flexible trimmer for Illumina sequence data\.*Bioinformatics***30**, 2114–2120 \(2014\)\. [Article](https://doi.org/10.1093%2Fbioinformatics%2Fbtu170)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2cXht1Sqt7nP)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=24695404)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4103590)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Trimmomatic%3A%20a%20flexible%20trimmer%20for%20Illumina%20sequence%20data&journal=Bioinformatics&doi=10.1093%2Fbioinformatics%2Fbtu170&volume=30&pages=2114-2120&publication_year=2014&author=Bolger%2CAM&author=Lohse%2CM&author=Usadel%2CB)
68. Dobin, A\. et al\. STAR: ultrafast universal RNA\-seq aligner\.*Bioinformatics***29**, 15–21 \(2013\)\. [Article](https://doi.org/10.1093%2Fbioinformatics%2Fbts635)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC38XhvV2gsbnF)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=23104886)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=STAR%3A%20ultrafast%20universal%20RNA-seq%20aligner&journal=Bioinformatics&doi=10.1093%2Fbioinformatics%2Fbts635&volume=29&pages=15-21&publication_year=2013&author=Dobin%2CA)
69. Liao, Y\., Smyth, G\. K\. & Shi, W\. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features\.*Bioinformatics***30**, 923–930 \(2014\)\. [Article](https://doi.org/10.1093%2Fbioinformatics%2Fbtt656)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2cXltFGqu7c%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=24227677)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=featureCounts%3A%20an%20efficient%20general%20purpose%20program%20for%20assigning%20sequence%20reads%20to%20genomic%20features&journal=Bioinformatics&doi=10.1093%2Fbioinformatics%2Fbtt656&volume=30&pages=923-930&publication_year=2014&author=Liao%2CY&author=Smyth%2CGK&author=Shi%2CW)
70. Jones, P\. et al\. InterProScan 5: genome\-scale protein function classification\.*Bioinformatics***30**, 1236–1240 \(2014\)\. [Article](https://doi.org/10.1093%2Fbioinformatics%2Fbtu031)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2cXmvFCjsr4%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=24451626)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3998142)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=InterProScan%205%3A%20genome-scale%20protein%20function%20classification&journal=Bioinformatics&doi=10.1093%2Fbioinformatics%2Fbtu031&volume=30&pages=1236-1240&publication_year=2014&author=Jones%2CP)
71. Kanehisa, M\., Sato, Y\. & Morishima, K\. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences\.*J\. Mol\. Biol\.***428**, 726–731 \(2016\)\. [Article](https://doi.org/10.1016%2Fj.jmb.2015.11.006)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2MXhvVOls7jI)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=26585406)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=BlastKOALA%20and%20GhostKOALA%3A%20KEGG%20Tools%20for%20Functional%20Characterization%20of%20Genome%20and%20Metagenome%20Sequences&journal=J.%20Mol.%20Biol.&doi=10.1016%2Fj.jmb.2015.11.006&volume=428&pages=726-731&publication_year=2016&author=Kanehisa%2CM&author=Sato%2CY&author=Morishima%2CK)
72. Grant, C\. E\., Bailey, T\. L\. & Noble, W\. S\. FIMO: scanning for occurrences of a given motif\.*Bioinformatics***27**, 1017–1018 \(2011\)\. [Article](https://doi.org/10.1093%2Fbioinformatics%2Fbtr064)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3MXkt1yqs74%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=21330290)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3065696)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=FIMO%3A%20scanning%20for%20occurrences%20of%20a%20given%20motif&journal=Bioinformatics&doi=10.1093%2Fbioinformatics%2Fbtr064&volume=27&pages=1017-1018&publication_year=2011&author=Grant%2CCE&author=Bailey%2CTL&author=Noble%2CWS)
73. Noon, J\. B\., Schwarz, E\. M\., Ostroff, G\. R\. & Aroian, R\. V\. A highly expressed intestinal cysteine protease of*Ancylostoma ceylanicum*protects vaccinated hamsters from hookworm infection\.*PLoS Negl\. Trop\. Dis\.***13**, e0007345 \(2019\)\. [Article](https://doi.org/10.1371%2Fjournal.pntd.0007345)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31009474)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6497320)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20highly%20expressed%20intestinal%20cysteine%20protease%20of%20Ancylostoma%20ceylanicum%20protects%20vaccinated%20hamsters%20from%20hookworm%20infection&journal=PLoS%20Negl.%20Trop.%20Dis.&doi=10.1371%2Fjournal.pntd.0007345&volume=13&publication_year=2019&author=Noon%2CJB&author=Schwarz%2CEM&author=Ostroff%2CGR&author=Aroian%2CRV)
74. Tintori, S\. C\., Sloat S\. A\., Rockman M\. V\. Rapid Isolation of Wild Nematodes by Baermann Funnel\.*J\. Vis\. Exp\.***179**,[https://doi\.org/10\.3791/63287](https://doi.org/10.3791/63287)\(2022\)\.
75. Clement, K\. et al\. CRISPResso2 provides accurate and rapid genome editing sequence analysis\.*Nat\. Biotechnol\.***37**, 224–226 \(2019\)\. [Article](https://doi.org/10.1038%2Fs41587-019-0032-3)[ADS](http://adsabs.harvard.edu/cgi-bin/nph-data_query?link_type=ABSTRACT&bibcode=2019NatBi..37..224C)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1MXosV2rtb0%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30809026)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6533916)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=CRISPResso2%20provides%20accurate%20and%20rapid%20genome%20editing%20sequence%20analysis&journal=Nat.%20Biotechnol.&doi=10.1038%2Fs41587-019-0032-3&volume=37&pages=224-226&publication_year=2019&author=Clement%2CK)
76. Magoc, T\. & Salzberg, S\. L\. FLASH: fast length adjustment of short reads to improve genome assemblies\.*Bioinformatics***27**, 2957–2963 \(2011\)\. [Article](https://doi.org/10.1093%2Fbioinformatics%2Fbtr507)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3MXhtlGkur7M)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=21903629)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3198573)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=FLASH%3A%20fast%20length%20adjustment%20of%20short%20reads%20to%20improve%20genome%20assemblies&journal=Bioinformatics&doi=10.1093%2Fbioinformatics%2Fbtr507&volume=27&pages=2957-2963&publication_year=2011&author=Magoc%2CT&author=Salzberg%2CSL)
77. Tedla, B\. A\. et al\. Novel cholinesterase paralogs of*Schistosoma mansoni*have perceived roles in cholinergic signalling and drug detoxification and are essential for parasite survival\.*PLoS Pathog\.***15**, e1008213 \(2019\)\. [Article](https://doi.org/10.1371%2Fjournal.ppat.1008213)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31809524)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6919630)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Novel%20cholinesterase%20paralogs%20of%20Schistosoma%20mansoni%20have%20perceived%20roles%20in%20cholinergic%20signalling%20and%20drug%20detoxification%20and%20are%20essential%20for%20parasite%20survival&journal=PLoS%20Pathog.&doi=10.1371%2Fjournal.ppat.1008213&volume=15&publication_year=2019&author=Tedla%2CBA)
78. Tedla, B\. A\., Pickering, D\., Becker, L\., Loukas, A\. & Pearson, M\. S\. Vaccination with Schistosoma mansoni Cholinesterases Reduces the Parasite Burden and Egg Viability in a Mouse Model of Schistosomiasis\.*Vaccines***8**, 162 \(2020\)\. [Article](https://doi.org/10.3390%2Fvaccines8020162)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3cXitlyitLvK)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32260125)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7349746)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Vaccination%20with%20Schistosoma%20mansoni%20Cholinesterases%20Reduces%20the%20Parasite%20Burden%20and%20Egg%20Viability%20in%20a%20Mouse%20Model%20of%20Schistosomiasis&journal=Vaccines&doi=10.3390%2Fvaccines8020162&volume=8&publication_year=2020&author=Tedla%2CBA&author=Pickering%2CD&author=Becker%2CL&author=Loukas%2CA&author=Pearson%2CMS)
79. Bae, D\. H\. et al\. Design and testing of vector\-producing HEK293T cells bearing a genomic deletion of the SV40 T antigen coding region\.*Mol\. Ther\. Methods Clin\. Dev\.***18**, 631–638 \(2020\)\. [Article](https://doi.org/10.1016%2Fj.omtm.2020.07.006)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3cXhvVeqsLfI)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32775497)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7397404)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Design%20and%20testing%20of%20vector-producing%20HEK293T%20cells%20bearing%20a%20genomic%20deletion%20of%20the%20SV40%20T%20antigen%20coding%20region&journal=Mol.%20Ther.%20Methods%20Clin.%20Dev.&doi=10.1016%2Fj.omtm.2020.07.006&volume=18&pages=631-638&publication_year=2020&author=Bae%2CDH)
80. Li, H\., Gazzola, D\., Hu, Y\. & Aroian, R\. V\. An efficient method for viable cryopreservation and recovery of hookworms and other gastrointestinal nematodes in the laboratory\.*Int J\. Parasitol\.***53**, 451–458 \(2023\)\. [Article](https://doi.org/10.1016%2Fj.ijpara.2023.05.001)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=37201563)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC10330584)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=An%20efficient%20method%20for%20viable%20cryopreservation%20and%20recovery%20of%20hookworms%20and%20other%20gastrointestinal%20nematodes%20in%20the%20laboratory&journal=Int%20J.%20Parasitol.&doi=10.1016%2Fj.ijpara.2023.05.001&volume=53&pages=451-458&publication_year=2023&author=Li%2CH&author=Gazzola%2CD&author=Hu%2CY&author=Aroian%2CRV)
81. Leinonen, R, Sugawara, H, Shumway, M, on behalf of the International Nucleotide Sequence Database C\. The Sequence Read Archive\.*Nucleic Acids Res\.***39**, D19–D21 \(2011\)\.
82. Sayers, E\. W\. et al\. GenBank 2025 update\.*Nucleic Acids Res\.***53**, D56–D61 \(2025\)\. [Article](https://doi.org/10.1093%2Fnar%2Fgkae1114)[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB28Xhs1eisbg%3D)[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=39558184)[PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC11701615)[Google Scholar](http://scholar.google.com/scholar_lookup?&title=GenBank%202025%20update&journal=Nucleic%20Acids%20Res.&doi=10.1093%2Fnar%2Fgkae1114&volume=53&pages=D56-D61&publication_year=2025&author=Sayers%2CEW)
[Download references](https://citation-needed.springer.com/v2/references/10.1038/s41467-026-73447-9?format=refman&flavour=references)
## Acknowledgements
We thank the Mass Spectrometry Technology Access Center at the McDonnell Genome Institute \(MTAC@MGI\) at Washington University School of Medicine for mass spectrometry data generation and the Genome Technology Access Center at the McDonnell Genome Institute \(GTAC @ MGI\) for RNAseq data generation and acquisition\.
## Funding
K\.S\.S\., S\.B\., B\.A\.R\., M\.B\., S\.C\.U\., Y\.J\.C\., J\.C\.M\., D\.K\., S\.P\.D\., D\.A\.P\., R\.R\., B\.K\.B\., M\.E\.B\., W\.I\., M\.Moyle, P\.J\.B\., A\.L\., S\.D\. and M\.M\. disclose support for the research of this work from the Defense Advanced Research Projects Agency \(DARPA\) and Naval Information Warfare Center Pacific \(NIWC Pacific\) \[Contract no\. N66001\-21\-C\-4013\]\. E\.C\.C\. discloses support for the research of this work from the Defense Advanced Research Projects Agency \(DARPA\) \[Contract no\. N660012314009\]\. The views, opinions, and/or findings expressed are those of the author and should not be interpreted as representing the official views or policies of the Department of Defense or the U\.S\. Government\. Distribution Statement “A” \(Approved for Public Release, Distribution Unlimited\)\.
## Author information
Author notes
1. These authors contributed equally: Kumar Sachin Singh, Suman Bharti, Bruce A\. Rosa\.
### Authors and Affiliations
1. Division of Infectious Diseases, Department of Medicine, Washington University School of Medicine, St\. Louis, MO, USA Kumar Sachin Singh, Suman Bharti, Bruce A\. Rosa, Mahdiyeh Bigham, Samuel C\. Uzoechi, Young\-Jun Choi, John C\. Martin & Makedonka Mitreva
2. Department of Cell Biology and Physiology, Washington University School of Medicine, St\. Louis, MO, USA Danielle Kemper, Slavica Pavlovic Djuranovic & Sergej Djuranovic
3. Department of Pathology and Laboratory Medicine, Brown University, Providence, RI, USA Slavica Pavlovic Djuranovic
4. Australian Institute of Tropical Health and Medicine, James Cook University, Cairns, QLD, Australia Darren A\. Pickering, Rachael Ryan, Matthew Moyle & Alex Loukas
5. Charles River Analytics, Inc, Cambridge, MA, USA Bethany K\. Bracken
6. National School of Tropical Medicine, Department of Pediatric Tropical Medicine, Baylor College of Medicine, Houston, TX, USA Maria Elena Bottazzi
7. Department of Environmental, Agricultural, and Occupational Health, College of Public Health, University of Nebraska Medical Center, Omaha, NE, USA Eric C\. Carnes
8. Department of Microbiology, Immunology & Tropical Medicine, School of Medicine and Health Sciences, George Washington University, Washington, DC, USA Wannaporn Ittiprasert & Paul J\. Brindley
9. College of Medicine and Dentistry, James Cook University, Cairns, QLD, Australia Alex Loukas
10. Department of Molecular Biology, Cell Biology and Biochemistry, Brown University, Providence, RI, USA Sergej Djuranovic
11. University of Montenegro, Institute for Interdisciplinary and Multidisciplinary Studies, Podgorica, Montenegro Sergej Djuranovic
12. Department of Genetics, Washington University School of Medicine, St\. Louis, MO, USA Makedonka Mitreva
13. McDonnell Genome Institute, Washington University School of Medicine, St\. Louis, MO, USA Makedonka Mitreva
Authors
1. Kumar Sachin Singh
2. Suman Bharti
3. Bruce A\. Rosa
4. Mahdiyeh Bigham
5. Samuel C\. Uzoechi
6. Young\-Jun Choi
7. John C\. Martin
8. Danielle Kemper
9. Slavica Pavlovic Djuranovic
10. Darren A\. Pickering
11. Rachael Ryan
12. Bethany K\. Bracken
13. Maria Elena Bottazzi
14. Eric C\. Carnes
15. Wannaporn Ittiprasert
16. Matthew Moyle
17. Paul J\. Brindley
18. Alex Loukas
19. Sergej Djuranovic
20. Makedonka Mitreva
### Contributions
Conceptualization: P\.J\.B\., A\.L\., S\.D\., M\.M\., Data curation: B\.A\.R\., Y\.J\.C\., J\.C\.M\., Formal Analysis: K\.S\.S\., S\.B\., B\.A\.R\., Y\.J\.C\., J\.C\.M\., D\.K\., S\.P\.D\., D\.A\.P\., R\.R\., E\.C\.C\., Funding acquisition: B\.K\.B\., P\.J\.B\., A\.L\., S\.D\., M\.M\., Investigation: B\.K\.B\., M\.E\.B\., M\.Moyle, P\.J\.B\., A\.L\., S\.D\., M\.M\., Methodology: K\.S\.S\., S\.B\., B\.A\.R\., M\.B\., S\.C\.U\., Y\.J\.C\., J\.C\.M\., D\.K\., S\.P\.D\., D\.A\.P\., R\.R\., E\.C\.C\., W\.I\., M\.Moyle, P\.J\.B\., A\.L\., S\.D\., M\.M\., Project administration: B\.K\.B\., P\.J\.B\., A\.L\., S\.D\., M\.M\., Resources: M\.E\.B\., E\.C\., W\.I\.T\., P\.J\.B\., A\.L\., S\.D\., M\.M\., Software: B\.A\.R\., Y\.J\.C\., J\.C\.M\., Supervision: P\.J\.B\., A\.L\., S\.D\., M\.M\., Validation: K\.S\.S\., S\.B\., W\.I\.T\., P\.J\.B\., A\.L\., S\.D\., M\.M\., Visualization: K\.S\.S\., S\.B\., B\.A\.R\., Writing \- original draft: K\.S\.S\., S\.B\., B\.A\.R\., M\.M\., Writing \- review & editing: K\.S\.S\., S\.B\., B\.A\.R\., M\.B\., S\.C\.U\., Y\.J\.C\., J\.C\.M\., D\.K\., S\.P\.D\., D\.A\.P\., R\.R\., B\.K\.B\., M\.E\.B\., E\.C\.C\., WI, M\.Moyle, P\.J\.B\., A\.L\., S\.D\., M\.M\.
### Corresponding author
Correspondence to[Makedonka Mitreva](mailto:
[email protected])\.
## Ethics declarations
### Competing interests
Makedonka Mitreva and Sergej Djuranovic are inventors on a patent related to the technology described in this manuscript \(US Patent Application Publication 20240424030\)\. The remaining authors declare no competing interests\.
## Peer review
### Peer review information
*Nature Communications*thanks the anonymous reviewers for their contribution to the peer review of this work\. A peer review file is available\.
## Additional information
**Publisher’s note**Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations\.
## Supplementary information
## Source data
## Rights and permissions
**Open Access**This article is licensed under a Creative Commons Attribution\-NonCommercial\-NoDerivatives 4\.0 International License, which permits any non\-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author\(s\) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material\. You do not have permission under this licence to share adapted material derived from this article or parts of it\. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material\. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder\. To view a copy of this licence, visit[http://creativecommons\.org/licenses/by\-nc\-nd/4\.0/](http://creativecommons.org/licenses/by-nc-nd/4.0/)\.
[Reprints and permissions](https://s100.copyright.com/AppDispatchServlet?title=Transgenic%20hookworm%20secretes%20anti-tetrodotoxin%20human%20single%20chain%20antibody&author=Kumar%20Sachin%20Singh%20et%20al&contentID=10.1038%2Fs41467-026-73447-9©right=The%20Author%28s%29&publication=2041-1723&publicationDate=2026-06-03&publisherName=SpringerNature&orderBeanReset=true&oa=CC%20BY-NC-ND)
## About this article
[](https://crossmark.crossref.org/dialog/?doi=10.1038/s41467-026-73447-9)
### Cite this article
Singh, K\.S\., Bharti, S\., Rosa, B\.A\.*et al\.*Transgenic hookworm secretes anti\-tetrodotoxin human single chain antibody\.*Nat Commun***17**, 4691 \(2026\)\. https://doi\.org/10\.1038/s41467\-026\-73447\-9
[Download citation](https://citation-needed.springer.com/v2/references/10.1038/s41467-026-73447-9?format=refman&flavour=citation)
- Received:17 January 2025
- Accepted:12 May 2026
- Published:03 June 2026
- Version of record:03 June 2026
- DOI:https://doi\.org/10\.1038/s41467\-026\-73447\-9